|
第三节 ATP的生成、储存和利用 ATP几乎是生物组织细胞能够直接利用的唯一能源,在糖、脂类及蛋白质等物质氧化分解中释放出的能量,相当大的一部分能使ADP磷酸化成为ATP,从而把能量保存在ATP分子内。 ATP为一游离核苷酸,由腺嘌呤、核糖与三分子磷酸构成,磷酸与磷酸间借磷酸酐键相连,当这种高能磷酸化合物水解时(磷酸酐键断裂)自由能变化(G)为30.5KJ/mol,而一般的磷酸酯水解时(磷酸酯键断裂)自由能的变化只有8至12KJ/mol,因此曾称此磷酸酐键为高能磷酸键,但实际上这样的名称是不够确切的,因为一种化合物水解时释放自由能的多少取决于该化合物整个分子的结构,以及反应的作用物自由能与产物自由能的差异,而不是由哪个特殊化学键的破坏所致,但为了叙述及解释问题方便,高能磷酸键的概念至今仍被生物化学界采用。 ATP是一高能磷酸化合物,当ATP水解时首先将其分子的一部分,如磷酸(Pi)或腺苷酸(AMP)转移给作用物,或与催化反应的酶形成共价结合的中间产物,以提高作用物或酶的自由能,最终被转移的AMP或Pi将被取代而放出,ATP多以这种通过磷酸基团等转移的方式,而非单独水解的方式,参加酶促反应提供能量,用以驱动需要加入自由能的吸能反应,ATP水解反应的总结如下: ATP——→ADP+Pi 或ATP——→AMP+PPi (焦磷酸) 一、ATP的生成方式 体内ATP生成有两种方式 (一)底物水平磷酸化(substrate level phosphorylation) 底物分子中的能量直接以高能键形式转移给ADP生成ATP,这个过程称为底物水平磷酸化,这一磷酸化过程在胞浆和线粒体中进行,包括有:
(二)氧化磷酸化(oxidative phosphorylation) 氧化和磷酸化是两个不同的概念。氧化是底物脱氢或失电子的过程,而磷酸化是指ADP与Pi合成ATP的过程。在结构完整的线粒体中氧化与磷酸化这两个过程是紧密地偶联在一起的,即氧化释放的能量用于ATP合成,这个过程就是氧化磷酸化,氧化是磷酸化的基础,而磷酸化是氧化的结果。 机体代谢过程中能量的主要来源是线粒体,既有氧化磷酸化,也有底物水平磷酸化,以前者为主要来源。胞液中底物水平磷酸化也能获得部分能量,实际上这是酵解过程的能量来源。对于酵解组织、红细胞和组织相对缺氧时的能量来源是十分重要的。 二、氧化磷酸化偶联部位的测定 确定氧化磷酸化偶联部位通常用两种方法。 (一)P/0值测定 P/0值指在氧化磷酸化过程中消耗一克原子氧所消耗的无机磷的克原子数,或者说消耗一克原子氧所生成的ATP的克分子数。P/0值实质上指的是呼吸过程中磷酸化的效率。 测定P/0值的方法通常是在一密闭的容器中加入氧化的底物、ADP、Pi、氧饱和的缓冲液,再加入线粒体制剂时就会有氧化磷酸化进行。反应终了时测定O2消耗量(可用氧电极法)和Pi消耗量(或ATP生成量)就可以计算出P/0值了。在反应系统中加入不同的底物,可测得各自的P/0值,结合我们所了解的呼吸链的传递顺序,就可以分析出大致的偶联部位了。? 表6-3 离体线粒体的P/0比值
从上表可以看出P/0值为小数,由于线粒体的偶联作用在离体条件下不能完全发挥,故可认为实际的ATP生成数是他们所接近的正整数值。 比较表中的(1)和(2),呼吸链传递的差异是在CoQ之间,两者ATP的生成数相差1,所以这个ATP的生成部位一定在NAD→CoQ之间。 比较表中(2)和(3),呼吸链传递的差异是在Cyt c之间,两者ATP的生成数相差1,所以这个ATP的生成部位在CoQ→Cyt c之间。 比较表中(3)和(4),生成的ATP数均为1,呼吸链传递的区别是在Cyt c→Cyt aa3,故Cytc→ aa3不存在偶联部位,而在Cyt aa3→O2之间存在着一个偶联部位。 (二)根据氧化还原电位计算电子传递释放的能量是否能满足ATP合成的需要 氧化还原反应中释放的自由能△G'O与反应底物和产物标准氧化还原电位差值(△E'O)之间存在下述关系:△G'O=?nF△E'O 式中n为氧化还原反应中电子转移数目,F为法拉弟常数(23.062千卡/克分子·伏特,或96500库仑/克分子)。 一克分子ATP水解生成ADP与Pi所释放的能量为7.3千卡,凡氧化过程中释放的能量大于7.3千,均有可能生成一克分子ATP,就是说可能存在有一个偶联部位,根据上式计算,当n=2时,△E'O=0.1583V时可释放7.3千卡能量,所以反应底物与生成物的标准氧化还原电位的变化大于0.1583V的部位均可能存在着一个偶联部位。 从图6-11可以看出,在NAD→CoQ,Cyt b→Cyt c和Cyt aa3→O2处可能存在着偶联部位。必须明确,这种计算的基础是反应处在热力学平衡状态,温度为25℃,pH为7.0,反应底物和产物的浓度均为1克分子,这种条件在体内是不存在的。因此这一计算结果只能供参考。
图6-11 呼吸链中电子对传递时自由能的变化 综上所述,呼吸链中电子传递和磷酸化的偶联部位可用图6-12表示。
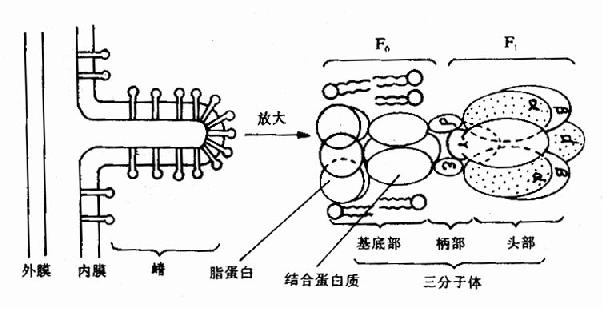
图6-12 偶联部位示意图 呼吸链磷酸化的全过程可用下述方程式表示 NADH+H++3ADP+3Pi+1/2O2→NAD++3ATP+4H2O FADH2+2ADP+2Pi+1/2O2→FAD+2ATP+3H2O 三、氧化磷酸化中ATP生成的结构基础 ATP是由位于线粒体内膜上的ATP合成酶催化ADP与Pi合成的。ATP合成酶是一个大的膜蛋白质复合体,分子量在480?00kD,是由两个主要组成(或称因子)构成,一是疏水的F0,另一是亲水的F1,又称F0F1复合体。在电子显微镜下观察线粒体时,可见到线粒体内膜基质侧有许多球状颗粒突起,这就是ATP合成酶,其中球状的头与茎是F1部分,分子量为350?/P>380kD,由α3、β3、γ、δ、ε等9种多肽亚基组成,β与α亚基上有ATP结合部位;γ亚基被认为具有控制质子通过的闸门作用;δ亚基是F1与膜相连所必需,其中中心部分为质子通路;ε亚基是酶的调节部分,F0是由3?个大小不一的亚基组成,其中有一个亚基称为寡霉素敏感蛋白质(oligomycin?sensitivity?conferringprotein,OSCP),此外尚有一个蛋白质部分为分子量28kD的因子,F0主要构成质子通道(见图6-13)。
图6-13 线粒体内膜上三联体(三分子体)结构示意图 四、氧化磷酸化的偶联机制 有关氧化磷酸化的偶联机理已经作了许多研究,目前氧化磷酸化的偶联机理还不完全清楚,50年代Slater及Lehninger提出了化学偶联学说,1964年Boear又提出了构象变化偶联学说,这两种学说的实验依据不多,支持这两种观点的人已经不多了。目前多数人支持化学渗透学说(chemiosmotic hypothesis),这是英国生化学家P.Mitchell于1961年提出的,当时没有引起人们的重视,1966年他根据逐步积累的实验证据和生物膜研究的进展,逐步地完善了这一学说。 氧化磷酸化的化学渗透学说的基本观点是: 1.线粒体的内膜中电子传递与线粒体释放H+是偶联的,即呼吸链在传递电子过程中释放出来的能量不断地将线粒体基质内的H+逆浓度梯度泵出线粒体内膜,这一过程的分子机理还不十分清楚(见图6-14)。
图6-14 电子传递与质子传递偶联(注:复合物Ⅱ未显示) 2.H+不能自由透过线粒体内膜,结果使得线粒体内膜外侧H+浓度增高,基质内H+浓度降低,在线粒体内膜两侧形成一个质子跨膜梯度,线粒体内膜外侧带正电荷,内膜内侧带负电荷,这就是跨膜电位△ψ。由于线粒体内膜两侧H+浓度不同,内膜两侧还有一个pH梯度△pH,膜外侧pH较基质pH约低1.0单位,底物氧化过程中释放的自由能就储存于△ψ和△pH中,若以△P表示总的质子移动力,那么三者的关系可用下式表示: △P=△ψ-59△pH 3.线粒体外的H+可以通过线粒体内膜上的三分子体顺着H+浓度梯度进入线粒体基质中,这相当于一个特异的质子通道,H+顺浓度梯度方向运动所释放的自由能用于ATP的合成,寡霉素能与OSCP结合,特异阻断这个H+通道,从而抑制ATP合成。有关ATP合成的分子机制目前还不十分清楚。 4.解偶联剂的作用是促进H+被动扩散通过线粒体内膜,即增强线粒体内膜对H+的通透性,解偶联剂能消除线粒体内膜两侧的质子梯度,所以不能再合成ATP。 总之,化学渗透学说认为在氧化与磷酸化之间起偶联作用的因素是H+的跨膜梯度。 每对H+通过三分子体回到线粒体基质中可以生成一分子ATP。以NADH+H+作底物,其电子沿呼吸链传递在线粒体内膜中形成三个回路,所以生成3分子ATP。以FADH2为底物,其电子沿琥珀酸氧化呼吸链传递在线粒体内膜中形成两个回路,所以生成两个ATP分子。 自从Mitchell提出化学通透学说以来,已为大量的实验结果验证,为该学说提供了实验依据。 美国Cohen等人于1978年使用完整的大鼠肝细胞作实验材料,以核磁共振(nuclear magneticresonance,NMR)的方法直接观察到完整细胞中胞液与线粒体基质之间存在H+跨膜梯度,胞液的pH值比线粒体基质的pH值低0.3单位,用解偶联剂处理,或用氮气代替氧气切断氧的供应,那么胞液和线粒体基质之间的pH梯度消失。 (责任编辑:泉水) |
第三节 ATP的生成、储存和利用
时间:2006-06-20 10:47来源:大众医药网 作者:admin 点击:
319次




顶一下
(4)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 发表评论
-
- 最新评论 进入详细评论页>>
