|
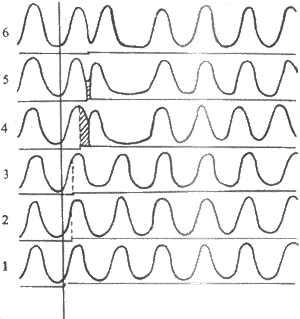
以下实验可以说明心肌组织的这一特点。正常情况下,窦房结产生的每一次兴奋传播到心房肌或心室肌的时间,都是在它们前一次兴奋的不应期终结之后,因此,整个心脏能够按照窦房结的节律而兴奋。但在某些情况下,如果心室在有效不应期之后受到人工的或窦房结之外的病理性异常异常刺激,则可产生一次期前兴奋,引起期前收缩或额外收缩。期前兴奋也有它自己的有效不应期,这样,当紧接在期前兴奋之后的一次窦房结兴奋传到心室肌时,常常正好落在期前兴奋的有效不应期内,因而不能引起心室兴奋和收缩,形成一次“脱失”,必须等到再下一次窦房结的兴奋传到心室时才能引起心室收缩。这样,在一次期前收缩之后往往出现一段较长的心室舒张期,称为代偿性间歇(图4-11)。随之,才恢复窦性节律。 (二)心肌的自动节律性 组织、细胞能够在没有外来刺激的条件下,自动地发生节律性兴奋的特性,称为自动节律性,简称自律性。具有自动节律性的组织或细胞,称自律组织或自律细胞。组织、细胞单位时间(每分钟)内能够自动发生兴奋的次数,即自动兴奋的频率,是衡量自动节律性高低的指标。
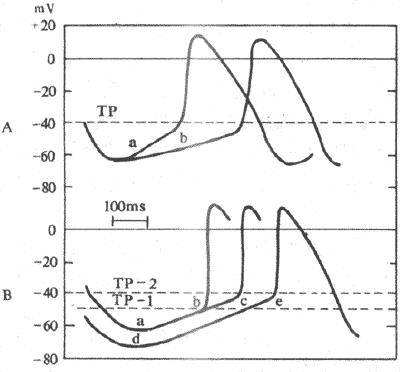
图4-11 期前收缩和代偿性间歇 每条曲线下的电磁标记号批示给予电刺激的时间,曲线1-3, 刺激落在有效不应期内,不引起反应;曲线4-6, 刺激落在相对不应期内,引起期前收缩和代偿性间歇 1.心肌的自动节律性和各自律级组织的相互关系很早以前就有人观察到,在适宜条件下,两栖类和哺乳类动物的离体心脏,在未受到任何刺激的情况下,可以长时间地、自动地、有节奏地进行兴奋和收缩。但是,只有到了近代,根据细胞内微电极技术记录的跨膜电位是否具有4期自动去极化这一特征,才确切地证明,并不是所有心肌细胞,而只是心脏特殊传导组织内某些自律细胞才具有自动节律性。特殊传导系统各个部位(结区除外)的自律性有等级差别;其中窦房结细胞自律性最高,自动兴奋频率约为每分钟100次,末梢浦肯野纤维网自律性最低(约每分钟25次),而房室交界(约每分钟50次)和房室束支的自律性依次介于两者之间。 由一个起搏点主宰整个心脏的整体活动具有极其重要的生理意义。那么,各部分自律组织的活动怎么能统一起来而不致于“各自为政”呢?实验中很容易观察到,心脏始终是依照当时情况下自律性最高的部位所发出的兴奋来进行活动的。这就是说,各部分的活动统一在自律性最高部位的主导作用之下。正常情况下,窦房结的自律性最高,它自动产生的兴奋向外扩布,依次激动心房肌、房室交界、房室束、心室内传导组织和心室肌,引起整个心脏兴奋和收缩。可见,窦房结是主导整个心脏兴奋和跳动的正常部位,故称为正常起搏点。其它部位自律组织并不表现出它们自身的自动节律性,只是起着兴奋传导作用,故称为潜在起搏点。在某种异常情况下,窦房结以外的自律组织(例如,它们的自律性增高,或者窦房结的兴奋因传导阻滞而不能控制某些自律组织)也可能自动发生兴奋,而心房或心室则依从当时情况下节律性最高部位的兴奋而跳动,这些异常的起搏部位则称为异位起搏点。 窦房结对于潜在起搏点的控制,通过两种方式实现:①抢先占领。窦房结的自律性高于其它潜在起搏点,所以,在潜在起搏点4期自动去极尚未达到阈电位水平之前,它们已经受到窦房结发出并依次传布而来的兴奋的激动作用而产生了动作电位,其自身的自动兴奋就不可能出现;②超速压抑或超速驱动压抑(overdrive suppression)。窦房结对于潜在起搏点,还可产生一种直接的抑制作用。例如,当窦房结对心室潜在起搏点的控制突然中断后,首先会出现一段时间的心室停搏,然后心室才能按其自身潜在起搏点的节律发生兴奋和搏动。出现这个现象的原因是:在自律性很高的窦房结的兴奋驱动下,潜在起搏点“被动”兴奋的频率远远超过它们本身的自动兴奋频率。潜在起搏长时间的“超速”兴奋的结果,出现了抑制效应;一旦窦房结的驱动中断,心室潜在起搏点需要一定的时间才能从被压抑状态中恢复过来,出现它本身的自动兴奋。另外还可以看到,超速压抑的程度与两个起搏点自动兴奋频率的差别呈平行关系,频率差别愈大,抑制效应愈强,驱动中断后,停搏的时间也愈长。因此,当窦房结兴奋停止或传导受阻后,首先由房室交界代替窦房结作为起搏点,而不是由心室传导组织首先代替;因为窦房结和房室交界的自动兴奋频率差距较小,超速压抑的程度较小。超速压抑产生的机制比较复杂,目前尚未完全弄清;但这一事实提示我们,在人工起搏的情况下,如因故需要暂时中断起搏器时,在中断之前其驱动频率应该逐步减慢,以避免发生心搏暂停。 2.决定和影响自律性的因素自律细胞的自动兴奋,是4期膜自动去极化使膜电位从最大复极电位达到阈电位水平而引起的。因此,自律性的高低,既受最大复极电位与阈电位的差距的影响,也取决于4期膜自动去极的速度(图4-12)。
图4-12 影响自律性的因素 A:起搏电位斜率由a减少到b时,自律性降低 B:最大复极电位水平由a达到d,或阈电位由TP-1升到TP-2时,自律性均降低 Tp :阈电位 (1)最大复极电位与阈电位之间的差距:最大复极电位绝对值减少和(或)阈电位下移,均使两者之间的差距减少,自动去极化达到阈电位水平所需时间缩短,自律性增高;反之亦然。例如,迷走神经系统兴奋时可使窦房结自律细胞K+通道开放率增高,故其复极3期内K+外流增加,最大复极电位绝对值增大,自律性降低,心率减慢。 (2)4期自动除极速度:4期自动除极速度与膜电位从最大复极电位水平达到阈电梯水平所需时间密切相关;若除极速度增快,达阈电位水平所需时间缩短,单位时间内发生兴奋的次数增多,自律性增高。从前一节已知,4期自动除极速度取决于净内向电流增长的速度,即取决于膜内净正电荷增长速度。例如,儿茶酚胺可以增强If,因而加速浦肯野细胞4期除极速度,提高其自律性。 (三)心肌的传导性和心脏内兴奋的传导 心肌在功能上是一种合胞体,心肌细胞膜的任何部位产生的兴奋不但可以沿整个细胞膜传播,并且可以通过闰盘传递到另一个心肌细胞,从而引起整块心肌的兴奋和收缩。动作电位沿细胞膜传播的速度可作为衡量传导性的指标。 1.心脏内兴奋传播的途径和特点 正常情况下窦房结发出的兴奋通过心房肌传播到整个右心房和左心房,尤其是沿着心房肌组成的“优势传导通路”迅速传到房室交界区,经房室束和左、右束支传到浦肯野纤维网,引起心室肌兴奋,再直接通过心室肌将兴奋由内膜侧向外膜侧心室肌扩布,引起整个心室兴奋。由于各种心肌细胞的传导性高低不等,兴奋在心脏各个部分传播的速度是不相同的。在心房,一般心房肌的传导速度较慢(约为0.4m/s),而“优势传导通路”的传导速度较快,窦房结的兴奋可以沿着这些通路很快传播到房室交界区。在心室,心室肌的传导速度约为1m/s,而心室内传导组织的传导性却高得多,末梢浦肯野纤维传导速度可达4m/s,而且它呈网状分布于心室壁,这样,由房室交界传入心室的兴奋就沿着高速传导的浦肯野纤维网迅速而广泛地向左右两侧心室壁传导。很明显,这种多方位的快速传导对于保持心室的同步收缩是十分重要的。房室交界区细胞的传导性很低,其中又以结区最低,传导速度仅0.02m/s。房室交界是正常时兴奋由心房进入心室的唯一通道,交界区这种缓慢传导使兴奋在这里延搁一段时间(称房-室延搁)才向心室传播,从而可以使心室在心房收缩完毕之后才开始收缩,不致于产生房室收缩重叠的现象。可以看出,心脏内兴奋传播途径的特点和传导速度的不一致性,对于心脏各部分有次序地、协调地进行收缩活动,具有十分重要的意义。 2.决定和影响传导性的因素 心肌的传导性取决于心肌细胞某些结构特点和电生理特性。 (1)结构因素:细胞直径与细胞内电阻呈反变关系,直径小的细胞内电阻大,产生的局部电流小于粗大的细胞,兴奋传导速度也较后者缓慢。心房肌、心室肌和浦肯野细胞的直径大于窦房结和房室交界细胞,其中,末梢浦肯野细胞的直径最大(在某些动物,直径可达70μm),兴奋传导速度最快;窦房结细胞直径很小(约5-10μm),传导速度很慢;而结区细胞直径更小,传导速度也最慢。 在机体生命过程中,心肌细胞直径不会突然发生明显的变化,因此,它只是决定传导性的一个比较固定的因素,对于各种生理或某些病理情况下心肌传导性的变化,不起重要作用。 (2)生理因素:心肌细胞的电生理特性是决定和影响心肌传导性的主要因素。与其它可兴奋细胞相同,心肌细胞兴奋的传播也是通过形成局部电流而实现的(参看第二章)。因此,可以从局部电流的形成和邻近未兴奋部位膜的兴奋性这两方面来分析影响传导性的因素。这两方面因素是密切相关联的,为了方便才分别叙述。 动作电位0期除极的速度和幅度局部电流是兴奋部位膜0期去极所引起的,0期去极的速度愈快,局部电流的形成也就愈快,很快就促使邻近未兴奋部位膜去极达到阈电位水平,故兴奋传导愈快。另一方面,0期去极幅度愈大,兴奋和未兴奋部位之间的电位差愈大,形成的局部电流愈强,兴奋传导也愈快。问题是,为什么局部电流的强度能影响传导速度?可能是强的局部电流扩布的距离大,可以使距兴奋部位更远的下游部位受到局部电流的刺激而兴奋,故兴奋的传导较快。除了细胞直径这个因素之外,浦肯野纤维等快反应细胞0期去极速度和幅度明显高于窦房结等慢反应细胞,是前者传导性比后者为高的主要原因。 已知,各种心肌细胞0期去极速度和幅度的差别,主要由膜上(0期)离子通道的固有性质决定。那么,同一心肌细胞(以快反应细胞为例)0期去极速度和幅度又受什么因素的影响?在叙述兴奋性时已经指出,快Na+通道的性状,即激活、失活和复活状态是决定兴奋性正常、缺失和低下的主要因素;也就是说,对兴奋性而言,Na+通道的性状决定着通道能否被激活开放(兴奋性的有无)以及激活的难易程度(兴奋性的高低)。这里,将进一步讨论,Na+通道的性状还决定着膜去极达阈电位水平后通道开放的速度和数量,从而决定膜0期去极的速度和幅度。Na+通道开放速度和数量这种性状,称为Na+通道的效率或可利用率(通道开放数量称开放概率)。实验证明,Na+通道的效率也是电压依从性的,它依从于临受刺激前的膜静息电位值。定量地分析Na+通道的效率(用0期去极的最大速率反映Na+通道开放的速度)与静息膜电位值的函数关系的曲线为膜反应曲线(图4-13)。膜反应曲线呈S形。正常静息电位值(-90mV)情况下,膜受刺激去极达阈电位水平后,Na+通道快速开放,0期去极最大速度可达500V/s。如膜静息电位值(绝对值)降低,去极最大速度下降;若膜静息电位值(绝对值)进一步降低到膜内为-60~-55mV时,去极速度几乎为0,即Na+通道已失活而不能开放。上述这种现象称为Na+通道效率的电压依从性下降。需要引起注意的是,在静息膜电位值(绝对值)很低(膜内-60~-55mV)状况下,如果膜受到刺激,并不是根本不产生电位变化,而是产生一种0期去极速度和幅度都很小的动作电位。这是因为,在这种情况下快Na+通道已经失活,而慢Ca2+通道未受影响,因此,原来的快反应细胞此时出现了由Ca2+内流所致的慢反应电位的缘故;兴奋传导速度也就明显减慢。不过,这已经是膜0期去极的离子通道发生了更换,不再属于Na+通道效率的量变范畴。 (责任编辑:泉水) |
第二节 心肌的生物电现象和生理特征(4)
时间:2006-06-19 20:38来源:大众医药网 作者:admin 点击:
777次
顶一下
(15)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 发表评论
-
- 最新评论 进入详细评论页>>
