|
平滑肌细胞中的细肌丝有同骨骼肌类似的分子结构,但不含肌钙蛋白;同一体积的平滑肌所含肌纤蛋白的量是骨骼肌的2倍,推测平滑肌肌浆中有大量细肌丝存在,它们的排列大致与细胞长轴平行。与此相反,胞浆中肌凝蛋白的量却只有骨骼肌的1/4。估计连接在致密体上的3~5根细肌丝会被较少数目粗肌丝包绕,形成相互交错式的排列,这可能就是类似于骨骼肌中肌小节的功能单位。
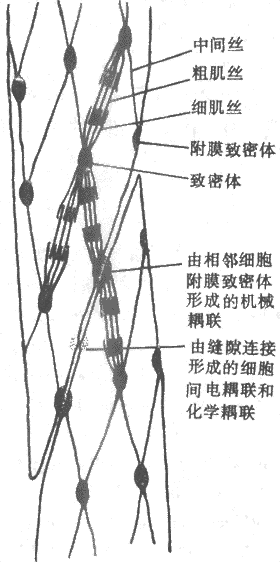
图2-29平骨肌内部结构和相互关系模式图 一般平滑肌细胞呈梭形,直径2~5μm;其长度可变性很大,大约长度为400μm时是产生张力的最适长度。它们没有骨骼肌(和心肌)那样发达的肌管系统。肌细胞膜只有一些纵向排列的袋状凹入,但其功能尚不清楚,不过这使得细胞膜表面积和细胞体积之比更为加大,因此和肌丝靠近的不是横管或肌浆网系统,而是肌膜。细胞被激活时,细胞外Ca2+进入膜内,但平滑肌细胞中靠近膜的肌浆网也构成了细胞内Ca2+贮存库。一些兴奋性递质、激素或药物同肌膜受体结合时,通过G-蛋白在胞浆中产生第二信使,引起Ca2+库中的Ca2+释出。因平滑肌的细肌丝中不存在肌钙蛋白,因而Ca2+引起平滑肌细胞中粗、细肌丝相互滑行的横桥循环的机制与骨骼肌不同。目前认为,横桥的激活开始于它的磷酸化,而这又依赖一称为肌凝蛋白激酶的活化;其过程是Ca2+先结合于胞浆中一种称为钙调蛋白(calmodulin)的特殊蛋白质,后者结合4个Ca2+之后才使肌凝蛋白激酶活化,使ATP分解,由此产生的磷酸基结合于横桥并使横桥处于高自由状态。比起平滑肌来,平滑肌横桥激活的机制需要较长的时间,这和平滑肌收缩的缓慢相一致。 (二)平滑肌在功能上的分类 尽管体内各器官所含平滑肌在功能特性上判别很大,但一般可分为两大类:一类称为多单位(multi-unit)平滑肌,其中所含各平滑肌细胞在活动时各自独立,类似骨骼肌细胞,如竖毛肌、虹膜肌、瞬膜肌(猫)、以及大血管平滑肌等,它们各细胞的活动受外来神经支配或受扩散到各细胞的激素的影响;另一类称为单位(single-unit)平滑肌,类似心肌组织,其中各细胞通过细胞间的电耦联而可以进行同步性活动,这类平滑肌大都具有自律性,在没有外来神经支配时也可进行近于正常的收缩活动(由于起搏细胞的自律性和内在神经丛的作用),以胃肠、子宫、输尿管平滑肌为代表。还有一些平滑肌兼有两方面的特点,很难归入哪一类,如小动脉和小静脉平滑肌一般认为属于多单位平滑肌,但又有自律性;膀胱平滑肌没有自律性,但在遇到牵拉时可作为一个整体起反应,故也列入单位平滑肌。 (三)平滑肌活动的控制和调节 和平滑肌本身的特性具有多样性一样,它们活动所受的调控也是多种多样的,不象骨骼肌那样单纯。大多数平滑肌接受神经支配,包括来自自主神经系统的外来神经支配,其中除小动脉一般只接受交感系统一种外来神经支配外,其他器官的平滑肌通常接受交感和副交感两种神经支配。平滑肌组织、特别是消化管平滑肌肌层中还有内在神经丛存在,后者接受外来神经的影响,但其中还发现有局部传入性神经元,可以引起各种反射。平滑肌的神经-肌接头有些类似骨骼肌,但不具有后者那样特殊结构形式。支配平滑肌的外来神经纤维在进入靶组织时多次分支,分支上每隔一定距离出现一个膨大,呈念珠状,称为曲张体,其中含有分泌囊泡,它们在神经冲动到达时可以释放其中递质或其他神经活性物质;每个曲张体和靶细胞的距离亦不固定,平均约为80~100nm,这说明由神经末梢释放出来的递质分子要扩散较远距离才能达到靶细胞,而靶细胞和神经末梢的关系也不可能是固定的;凡是递质分子可以到达而又具有该递质受体的平滑肌细胞,都可能接受外来神经的影响。 平滑肌细胞约保持-55~-60mV的静息电位,产生机制和骨骼肌类似。单位平滑肌细胞有产生动作电位的能力,而且通过细胞间通道可使相邻细胞也产生动作电位。 参考资料 |
第四节 肌细胞的收缩功能(4)
时间:2006-06-19 20:38来源:大众医药网 作者:admin 点击:
717次
顶一下
(13)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 发表评论
-
- 最新评论 进入详细评论页>>
