细胞信号转导与疾病 > 第一节 细胞信号转导的主要途径
第一节 细胞信号转导的主要途径(The major pathways of cellular signal transduction )
一、G蛋白介导的细胞信号转导途径(G pretein-mediated signal transduction)
G蛋白 是指可与鸟嘌呤核苷酸可逆性结合的蛋白质家族
(一) 腺苷酸环化酶途径(Adenylyl Cyclase Signal Transduction Pathway)
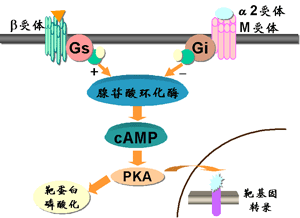
在腺苷酸环化酶(adenylyl cyclase,AC)信号转导途径中存在着两种作用相反的G蛋白,Gs与Gi。它们通过增加或抑制AC活性来调节细胞内cAMP浓度,进而影响细胞的功能。β肾上腺素受体、胰高血糖素受体等激活后经Gs增加AC活性,促进cAMP生成。而α2肾上腺素能受体、M2胆碱能受体及血管紧张素II受体等激活则与Gi偶联,经抑制AC活性减少cAMP的生成。cAMP可激活蛋白激酶A(protein kinase A,PKA),引起多种靶蛋白磷酸化,调节其功能。例如,肾上腺素引起肝细胞内cAMP增加,通过PKA促进磷酸化酶激酶活化,增加糖原分解。心肌β受体兴奋引起的cAMP增加经PKA促进心肌钙转运,提高心肌收缩力。进入核内的PKA可磷酸化转录因子CRE结合蛋白(cAMP response element binding protein, CREB),使其与DNA调控区的cAMP反应元件(cAMP response element, CRE)相结合,激活靶基因转录(图13-1)。
图13-1 腺苷酸环化酶信号转导通路
(二)IP3 、Ca2+ -钙调蛋白激酶途径
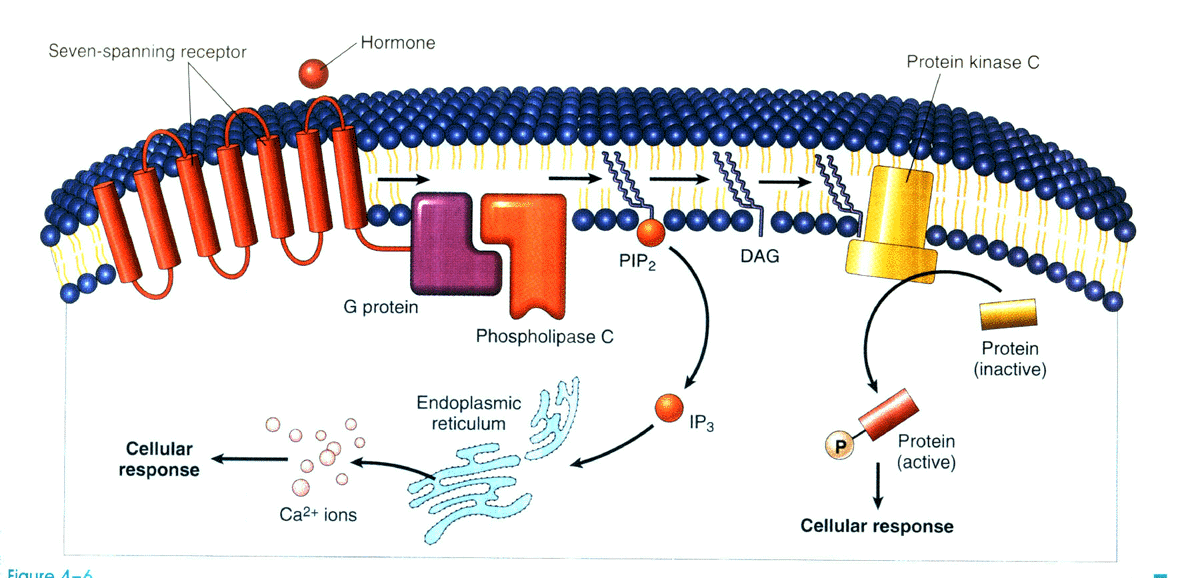
α1肾上腺素能受体、内皮素受体、血管紧张素II受体等激活可与Gqα结合,激活细胞膜上的磷脂酶C(phospholipase C, PLC)β亚型,催化质膜磷脂酰肌醇二磷酸(phosphatidylinositol 4,5-diphosphate,PIP2 )水解,生成三磷酸肌醇(1,4,5-inositol triphosphate, IP3 )和甘油二酯(1,2-diacylglycerol,DG)。IP3 促进肌浆网或内质网储存的Ca2+ 释放,Ca2+ 亦可作为第二信使启动多种细胞反应。例如,促进胰岛β细胞释放胰岛素;与心肌和骨骼肌的肌钙蛋白结合,触发肌肉收缩。Ca2+ 与钙调蛋白结合,激活Ca2+ /钙调蛋白依赖性蛋白激酶活性,经磷酸化多种靶蛋白产生生物学作用。
(三)DG-蛋白激酶C途径
DG与Ca2+ 能协调促进蛋白激酶C(protein kinase C,PKC)活化。激活的PKC可促进细胞膜Na+ /H+ 交换蛋白磷酸化,增加H+ 外流;PKC激活也可通过磷酸化转录因子AP-1、NF- KB 等,促进靶基因转录和细胞的增殖与肥大(图13-2)。
二、酪氨酸蛋白激酶介导的信号转导途径(Tyrosine Protein Kinase-mediated Signal Transduction Pathway)
(一)受体酪氨酸蛋白激酶途径(Receptor Tyrosine Protein Kinase Pathway)
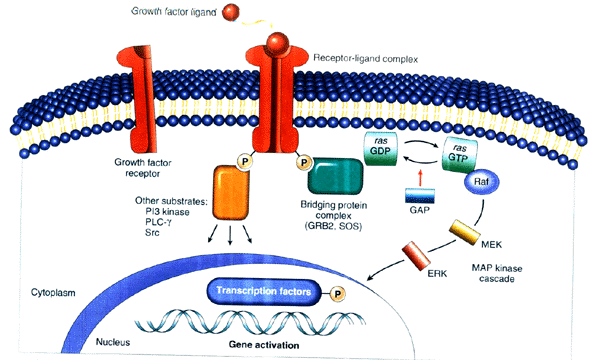
受体酪氨酸蛋白激酶(tyrosine protein kinase,TPK)是由50多种跨膜受体组成的超家族,其共同特征是受体胞内区含有TPK,配体则以生长因子为代表。表皮生长因子(epidermal growth factor, EGF)、血小板源生长因子(platelet-derived growth factor, PDGF)等与受体胞外区结合后,受体发生二聚化并催化胞内区酪氨酸残基自身磷酸化,进而活化TPK。磷酸化的酪氨酸可被一类含有SH2区(Src homology 2 domain)的蛋白质识别,通过级联反应向细胞内进行信号转导。由于大多数调节细胞增殖及分化的因子都通过这条途径发挥作用,故它与细胞增殖肥大和肿瘤发生的关系十分密切。
1.经Ras蛋白激活丝裂原活化蛋白激酶
丝裂原活化蛋白激酶(mitogen activited protein kinase,MAPK)家族是与细胞生长、分化、凋亡等密切相关的信号转导途径中的关键物质,可由多种方式激活。EGF、PDGF等生长因子与其受体结合并引起TPK激活后,细胞内含SH2区的生长因子受体连接蛋白Grb2与受体结合,将胞浆中具有鸟苷酸交换因子活性的Sos吸引至细胞膜,Sos 促进无活性Ras所结合的GDP为GTP 所置换,导致Ras活化。激活的Ras活化Raf(又称MAPK kinase kinase,MAPKKK),进而激活MEK(又称MAPK kinase,MAPKK),最终导致细胞外信号调节激酶(extracellular signal regulated kinase,ERK)激活(图13-3)。激活的ERK可促进胞浆靶蛋白磷酸化或调节其他蛋白激酶的活性,如激活磷脂酶A2;激活调节蛋白质翻译的激酶等。更重要的是激活的ERK进入核内,促进多种转录因子磷酸化,如ERK促进血清反应因子(serum response factor, SRF)磷酸化,使其与含有血清反应元件(serum response element, SRE)的靶基因启动子相结合,增强转录活性。
2.经磷脂酶Cγ激活蛋白激酶C
受体TPK的磷酸化酪氨酸位点可为含有SH2区的PLCγ结合,导致PLCγ激活,水解PIP2 生成IP3 和DG,进而调节细胞的活动。
3.激活磷脂酰肌醇3激酶(phosphoinsitol 3' kinase,PI3 K)
PI3 K是由 p85调节亚单位和p110催化亚单位组成的异二聚体,因可催化磷脂酰肌醇3位的磷酸化而得名。PI3 K的 p85与受体磷酸化的酪氨酸相结合,调节p110催化亚单位的活性,促进底物蛋白磷酸化,在细胞生长与代谢的调节中发挥重要作用。例如,PI3 K可促进细胞由G1期进入S期;p110能与Ras-GTP结合,参与细胞生长的调节。
(二)非受体酪氨酸蛋白激酶信号转导途径
细胞因子如白介素(IL)、干扰素(INF)及红细胞生成素等的膜受体本身并无蛋白激酶活性,其信号转导是由非受体TPK介导的。非受体TPK的调节机制差异较大,现以INFγ为例,说明其信号转导途径。INFγ与受体结合并使受体发生二聚化后,受体的胞内近膜区可与胞浆内非受体TPK JAK激酶(janus kinase)结合并发生磷酸化,进而与信号转导和转录激活因子(signal transducers and activators of transcription,STAT)相结合。在JAK催化下,STAT中的酪氨酸磷酸化,并结合成STAT二聚体转移入核,与DNA启动子的活化序列结合,诱导靶基因的表达,促进多种蛋白质的合成,进而增强细胞抵御病毒感染的能力。
三、鸟苷酸环化酶信号转导途径
鸟苷酸环化酶(guanylyl cyclase,GC)信号转导途径存在于心血管系统和脑内,一氧化氮(nitric oxide,NO)激活胞浆可溶性GC,心钠素及脑钠素激活膜颗粒性GC,增加cGMP生成,再经激活蛋白激酶G(protein kinase G,PKG)磷酸化靶蛋白发挥生物学作用。
四、核受体(Nuclear receptor)及其信号转导途径
细胞内受体分布于胞浆或核内,本质上都是配体调控的转录因子,均在核内启动信号转导并影响基因转录,故统称为核受体(nuclear receptor) 。按核受体的结构与功能可将其分为:①类固醇激素受体家族 包括糖皮质激素、盐皮质激素、性激素受体等。类固醇激素受体(除雌激素受体位于核内)位于胞浆,未与配体结合前与热休克蛋白(heat shock protein,HSP)结合存在,处于非活化状态。配体与受体的结合使HSP与受体解离,暴露DNA结合区。激活的受体二聚化并转移入核,与DNA上的激素反应元件(hormone response element,HRE)相结合或与其他转录因子相互作用,增强或抑制靶基因转录;②甲状腺素受体家族 包括甲状腺素、维生素D和维甲酸受体等。此类受体位于核内,不与HSP结合,多以同源或异源二聚体的形式与DNA或其他蛋白质结合,配体入核与受体结合后,激活受体并经HRE调节基因转录。
近年来的研究表明,不同信息分子、不同信号转导途径之间还存在交互调节(cross-talk),从而构成复杂的信号转导网络。例如,Giβγ可通过激活PLCβ,引起AC及PLC介导的信号转导途径之间的交互调节;某些GPCRs可通过促进非受体TPK磷酸化,增加Grb2和Sos结合,以Ras依赖形式激活ERK;或是经PKC激活Raf,以非Ras依赖形式激活ERK。在众多的信号转导途径中,无论任何环节出现异常,都可使相应的信号转导过程受阻,导致细胞的应答反应减弱、丧失或者反应过度,这些均可导致疾病的发生。
(责任编辑:泉水)