哺乳动物神经元培养细胞中的RNAi作用
Anna M. Krichevsky* and Kenneth S. Kosik
Center for Neurologic Diseases, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA 02115
Edited by Solomon H. Snyder, Johns Hopkins University School of Medicine, Baltimore, MD,
and approved July 8, 2002 (received for review May 7, 2002)
在众多有机体中,双链RNA能触发转录后基因沉默或RNA干扰(RNA interference,RNAi)。这种生理途径的媒介,小的干扰RNA:21nt的双链RNA,已经成为敲除特定基因在哺乳动物细胞系中表达的强大工具,对分析功能显性缺陷型也非常有用。在基因操作非常困难的哺乳动物原代神经元培养物,RNAi很可能发展成为研究特定基因在神经元发育及功能中的高效工具。而此前,神经元曾被认为最能抵抗RNAi。我们在这儿报道RNAi在有丝分裂期后的原代神经元培养物中的应用:合成的siRNA能导入神经元并有效地抑制内源及转基因的表达。
双链RNA(dsRNA)通过RNAi或序列特异性转录后基因剪切途径(1-3)有效地抑制线虫(Caenorhabditis elegans), 果蝇(Drosophila melanogaster), 锥体虫(Trypanosoma bruce)及植物中特定基因表达。DsRNA被ribonuclease III剪切为21-22nt的RNA双链或小的干扰RNA(siRNA)。这些分子触发同源mRNA的降解(4,5)。尽管dsRNA导致哺乳动物细胞中整体性的蛋白合成关闭,如果直接导入小于30nt的siRNA就能抑制哺乳动物细胞系中特定的内源和异源基因的表达(6,7)。从这些研究中得出的结论是RNA干扰能够在哺乳动物细胞中发生,但siRNA触发的RNAi机制还有待阐明。现存的一些抑制哺乳细胞特定基因表达的方法(mainly antisense and dominant–negative)的效果和重复性都不理想。而RNAi,由于能在极低浓度,微量(nmol范围)siRNA存在情况下显示出特殊有效性,在研究哺乳动物细胞培养物的基因功能方面有极大的应用前景。许多基因已被成功地从哺乳动物体细胞和胚胎细胞系中敲除,包括常用的HeLa, HEK293, 和 P19细胞(8,9)。但由于一些尚不清楚的原因,神经元比其它细胞类型显示出对RNAi更强的抵抗力,可能这些细胞中RNA进入细胞膜或RNAi途径方面不太一样。在线虫(C. elegans)中,尽管通过喂食或注射dsRNA到整个动物体诱导RNAi的方法已经比较系统,甚至能抑制子代的基因表达,虫体的神经元却表现出RNAi抗性(10)。但,高浓度的dsRNA(15ug/ml)能够抑制增殖和分化的线虫神经元培养物靶基因的表达(11)。在D. melanogaster中,基因组cDNA杂交链(推测它能用于合成dsRNA)能作用于神经元中的靶基因表达(12)。本文中,我们将阐述导入极低浓度的siRNA到从大鼠海马和前脑中制备的解离的有丝分裂后培养物中,能有效抑制其内源和异源基因表达。
材料和方法
皮层和海马(Hippocampal)细胞培养物。
怀有18天胚胎(E18)的Sprague–Dawley大鼠通过注射CO2杀死,剖腹,迅速取出胚胎。取出大脑皮层和海马,37°C下,在含0.25%胰岛素的HBSS(Hepes-buffered Hanks’ balanced salt solution)溶液中消化15分钟。用HBSS洗组织三次,在火边,用巴斯德吸管(Pasteur pipette)手工解离。在含DMEM及5%(vol/vol)的FBS种植培养基中,将细胞以12000-20000/cm2的浓度种植在多聚赖氨酸包被的盖玻片上。3h后,将培养基换为Neurobasal,含B27添加剂和0.5mM谷氨酸盐。所有实验在细胞种植后5-8天开始。在这些培养物中几乎观察不到神经胶质细胞。
siRNA制备和转染。
pEGFP或dsRed2报告基因和MAP2或YB-1 mRNA相应的siRNA根据文献(13)推荐,每条链的5’端为磷酸盐,3’羟基化,有两个碱基外悬,通过化学合成而得。下列为实验成功的基因特异性序列:si-GFP正链:5’- CAAGCUGACCCUGAAGUUCUU-3’反链:5’ -GAACUUCAGGGUCAGCUUGUG-3’;si-DsRed正链:5’ -AGUUCCAGUACGGCUCCAAUU-3’反链:5’ -UUGGAGCCGUACUGGAACUUG-3’MAP2 siRNA1正链:5’ -CGAGAGGAAAGACGAAGGAUU-3’反链:5’ -UCCUUCGUCUUUCCUCUCGUG-3’siRNA2正链:5’ -CAGGGCACCUAUUCAGAUAUU-3’反链:5’ -UAUCUGAAUAGGUGCCCUGUG-3’形成双链siRNA的退火处理参见参考文献(6)描述。
报告质粒和siRNA的共转染用转染试剂Lipofectamine 2000 (Life Technologies)转染试剂在6孔板中进行。先用Opti-MEM稀释Lipofectamine,加到质粒/dsRNA中形成混合物,这个过程需25分钟。每孔加1ug pEGFP-C2,1 ug pDsRed2, 及 0.01–0.2 ug 21-bp dsRNA (siRNA),与2ul Lipofectamine形成终体积为1.7ml的混合液。转染2小时后换上Neurobasal,细胞固定后培养后24-48小时分析结果。转染效率通过1% 到8%的4’,6-diamidino-2- phenylindole染色检测。
内源靶基因的siRNA的转染用TransMessenger转染试剂(Qiagen, Chatsworth, CA)。根据厂家说明书,siRNA(每孔1ug)通过Enhanser R试剂浓缩并与4ul TransMessenger形成复合物。转染复合物稀释到900ulNeurobasal中,与细胞混合;2小时后它被Neurobasal替代。转染48-68小时后染细胞分析。分析结果通过三个独立的实验证实。
免疫细胞化学。
对MAP2和几种蛋白进行双标记免疫荧光标记。生长在玻璃盖玻片上的神经元用PBS洗,4%(w/vol)多聚甲醛固定,0.25% Triton X-100穿透,再用PBS洗两次以上,用10%(vol/vol)山羊血清封闭。细胞用一抗 (1:200稀释到含1%山羊血清的PBS中)4°C敷育过夜,PBS洗三次。MAP2的小鼠单抗由实验室自制(14),YB1的兔多抗由V. Evdokimova (McGill Univ., Montreal, QC, Canada)馈赠;cortactin的兔多抗由Santa Cruz Biotechnology公司提供。MAP2通过Alexa 488或 Alexa 594荧光偶连的山羊抗小鼠的二抗(Molecular Probes公司)检测。其他抗体通过488- 或Alexa 594-偶连的山羊抗兔的二抗(1:100; Molecular Probes)检测。最后,细胞用PBS洗,用抗萃灭的封片剂在显微镜下封片。
显微镜和成像分析。
荧光显微镜结合配套的共聚焦激光扫描系统(Zeiss Axiovert S100)检测结果。在相等曝光时间下记录所有特定报告质粒或特定抗体成像结果(非饱和条件)。每个共转染实验,至少随机选择分析了50个的转染神经元。针对MAP2靶蛋白的实验,每种实验条件至少分析了70个随机选择的神经元,对对照和靶细胞的基因表达都进行了定量。通过AdobePHOTOSHOP软件对像素密度定量,并进行扣除背景标准化处理。
结果
首先,为验证培养神经元RNAi的有效性,针对EGFP-C2和DsRed2报告载体的21nt正义和反义ssRNA选自编码区,具体设计和描述见材料和方法。每条链分别合成,退火形成具有siRNA特点的dsRNA(13)。原代神经元种植后5-8天转入这两种质粒及相应dsRNA。转染后两小时,培养基换为Neurobasal;固定细胞,24-48小时后观测EGFP和DsRed2的表达。荧光显微镜观察,几乎所有转染的神经元中EGFP或DsRed2表达明显被相应dsRNA特异性抑制,而不被无关dsRNA抑制。共转染这两种质粒24小时后大多数转染的神经元(效率1-8%)高水平表达GFP,DsRed水平略低一些,混在一起呈现出黄绿色。针对绿色荧光蛋白的siRNA(siGFP)明显降低了增强型绿色荧光蛋白(EGFP)的表达,而对dsRed表达无影响,因此细胞呈现红色。为了量化siRNA对GFP表达的影响,对特定的报告质粒在非饱和条件下记录曝光时间相等的影像,并随机选择了转染细胞作进一步的绿色和红色荧光检测。对每个细胞,绿色和红色荧光都进行了扣除背景的标准化处理。每次实验,至少比较了50个随机选择的转染对照和靶神经元。这些分析表明GFP表达被相应dsRNA特异性抑制,在红色荧光强度不变情况下,检测到的绿色荧光强度下降了42%。对每个分析的细胞,红色和绿色荧光的比例计算为arctangent值;结果分布如图B。分布的迁移代表siGFP对GFP表达的特异性抑制。此外,对照和靶细胞几乎没有重叠分布清楚地表明siGFP即便浓度最低(6ng/ml)也能影响几乎所有转染神经元。这是dsRNA的特异性抑制,无论正链和反义链ssRNA都不能抑制报告基因的表达图C.

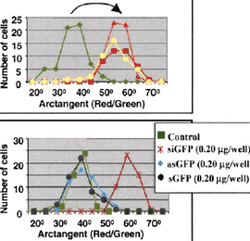
同样的对DsRed2与相应dsRNA的分析表明对照与靶细胞有一些重叠,抑制了约30%的DsRed2的表达。这由于后期表达的DsRed2的影响以及它在不同对照细胞中表达水平存在较大的差异所致。
我们还检测了siRNA对微管结合蛋白2(MAP2)和RNA结合蛋白YB-1(也称为p50)内源性mRNA的作用。在这些实验中,我们注意到siRNA相对于报告载体能很容易地导入神经元。因此无需导入报告基因作为细胞转入siRNA的标记。针对MAP2内源性mRNA的相应21nt dsRNA通过TransMessenger转染试剂转染细胞。转染后48-68小时,双免疫荧光标记MAP2和三个无关蛋白(cortactin,肌动蛋白,YB-1)。在70-80%的细胞中MAP2表达被相应的dsRNA特异性抑制,而无关蛋白的表达不受影响。在这些细胞中,MAP2荧光强度降低了4倍。在总的细胞中,由于转染效率及神经元对siRNA的敏感度不同荧光强度变化很大。不过,就从总细胞的统计荧光强度来说,MAP2荧光的降低还是很容易检测到。非常明显的是,统计分析说明两种针对MAP2的不同siRNA(siRNA1和siRNA2)对MAP2有同等的抑制作用。
MAP2抑制在培养神经元中的表型影响可通过神经元种植后纤维加固,出现许多丝状伪足等早期观察而得。已经有文献(15,16)表明MAP2是神经炎从早期进一步发展所必须的。我们用相应siRNA处理种植3小时后的原代神经元,在28-40小时左右分析结果。在这个短一些的时间段中,神经元MAP2表达抑制极少。但,MAP2抑制水平与观察到的丝状伪足缺乏程度一致。转染针对YB-1 RNA结合蛋白的不同dsRNA,即使敷育68小时条件下也不能抑制靶蛋白。这种情况下RNAi无效的原因可能由于YB-1蛋白丰度很高,又非常稳定或YB-1mRNA的特殊性质。
讨论
小的干扰RNA已经作为敲除特定基因表达的强大工具在很多细胞中运用。在果蝇裂解液中,dsRNA触发识别和靶向性剪切同源细胞mRNA(4)。在哺乳动物细胞中,siRNA介导的基因导向机制还不明确;可能影响RNA或翻译或转录稳定性。在培养的哺乳动物细胞中,化学合成的siRNA能通过亲脂性转染试剂导入细胞。最近,许多研究组发展了另一个通过转染质粒DNA胞内表达siRNA抑制基因表达的方法(17-20)。众所周知,转染有丝分裂后期的原代神经元效率非常低,通常有毒性,因此还没有报道过如本文所述的对神经元有效的siRNA介导的基因靶向性技术。原则上,RNA能通过阳离子脂类导入单个的神经元(21)。我们已经阐明合成的21nt dsRNA(siRNA)很容易通过阳离子的TransMessenger转染试剂导入到原代皮层和海马神经元中。因此,RNAi途径看来在原代哺乳动物神经元中是可行的。即使在简单敷育(2小时)在低浓度siRNA(0.006-0.6ug/ml)下也能导致内源和外源基因表达的抑制。同样浓度的反义ssRNA不能抑制基因表达。更高浓度的siRNA/转染试剂看来对神经元培养物有毒性。siRNA对MAP2表达的作用较慢,在转染后两天逐渐可观测到。我们推测更好的抑制转入的GFP和DsRed基因需要更长的细胞敷育时间;但我们用Lipofectamine 2000转染超过48小时后,神经元不能保持正常状态。除此之外唯一的RNAi有效的神经元培养系统,线虫类, RNAi作用也非常慢(dsRNA处理后3-4天不等),并需要量大得多的dsRNA(15ug/ml,而不是0.006-0.6ug/ml)。
目前,我们不知道是什么因素决定RNAi诱导的转录后基因抑制作用的速率和强度。比较清楚的是,靶mRNA(导致一些RNAi失效的“防卫性”二级结构和结合的转录因子)和蛋白(不同的生命周期)的特性影响RNAi效果。此外,细胞特异性因子可能参与不同细胞类型中不同siRNA运输和RNA干扰的调节。近来发现了一种以前没有明确分类的非编码RNA,主要分布在脑中的特异性微小RNA(microRNA)与siRNA共享一个生理途径(22),结合我们的数据显示RNAi相关机制在神经元发育和功能中对基因调控方面的作用。在基因操作非常困难的神经元中,敲除特定基因表达的技术方法将有广泛的应用。