|
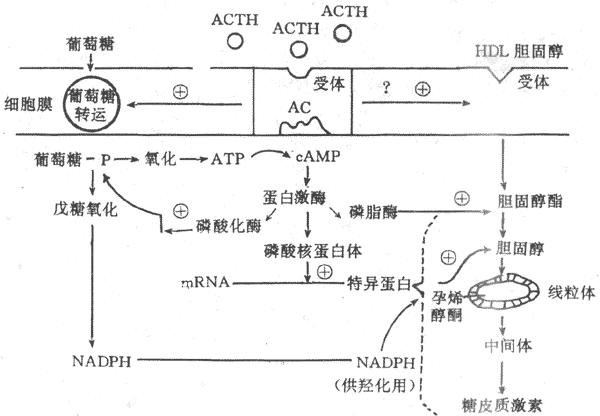
ACTH的分泌呈现日节律波动,入睡后ACTH分泌逐渐减少,午夜最低,随后又逐渐增多,至觉醒起床前进入分泌高峰,白天维持在较低水平,入睡时再减少。由于ACTH分泌的日节律波动,促糖皮质激素的分泌也出现相应的波动。ACTH分泌的这种日节律波动,是由下丘脑CRH节律性释放所决定的。 ACTH 不但刺激糖皮质激素的分泌,也刺激束状带与网状带细胞的生长发育,关于ACTH的作用机制已基本清楚。在束状还与网状带细胞膜上存在ACTH特异性受体,在Ca2+存在的条件下,ACTH与膜受体结合,激活腺苷酸环化酶,通过cAMP激活蛋白激酶,蛋白激酶起三项重要作用;①使核糖蛋白磷酸化,促进mRNA形成一种特殊蛋白质,使胆固醇得以进入线粒体,并经侧链解形成孕烯醇酮,以进一步合成糖皮质激素;②使磷酸化酶活化,促进糖原分解,产生ATP,提供能量,另外还通过戊糖旁路产生还原型辅酶Ⅱ(NADPH),以利胆固醇的羟化过程;③使胆固醇酯活化,促进其转变为胆固醇,提供激素合成的原料。在ACTH促进肾上腺皮质细胞合成糖皮质激素的同时,束状带细胞膜对葡萄糖与胆固醇的转运机制增强,使较多的葡萄糖与胆固醇进入细胞内(图11-18)。
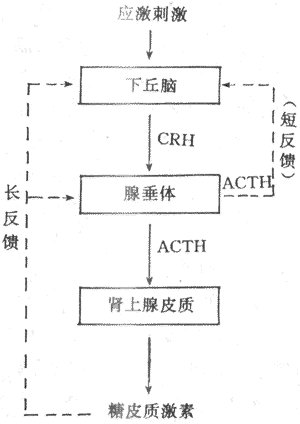
图11-18 ACTH作用机制示意图 HDL:高密度脂蛋白 AC:腺苷酸环化酶 (2)ACTH分泌的调节:ACTH调节糖皮质激素的分泌,而ACTH的分泌受下丘脑CRH的控制又与糖皮质激素有反馈调节。下丘脑CRH神经元和其他下丘脑调节肽神经元一样,又受脑内神经递质的调控。应激刺激作用于神经系统的不同部位,最后通过神经递质,将信息汇集于CRH神经元,然后借CRH控制腺垂体的促肾上腺皮质激素细胞分泌ACTH。此外,当血中糖皮质激素浓度升高时,可使腺垂体释放ACTH减少,ACTH的合成也受到抑制,腺垂体对CRH的反应也性减弱。糖皮质激素的负反馈调节主要作用于垂体,也可作用于下丘脑,这后一种反馈称为长反馈。ACTH还可反馈抑制CRH神经元,称为短反馈。至于是否存在CRH对CRH神经地的超短反馈,尚不能肯定。 综上所述,下丘脑、垂体和肾上腺皮质组成一个密切联系、协调统一的功能活动轴,从而维持血中糖皮质激素浓度的相对稳定和在不同状态下的适应性变化(图11-19)。 2.盐皮质激素分泌的调节 醛固酮的分泌主要受肾素-血管紧张素系统的调节。另外,血K+,血Na+浓度可以直接作用于球状带,影响醛固酮的分泌(详见第四章与第八章)。
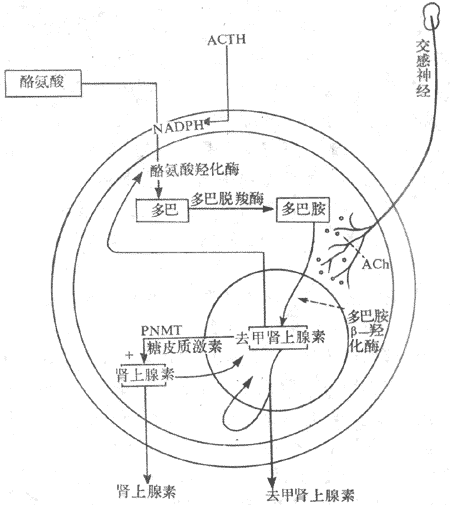
图11-19 糖皮质激素分泌的调节示意图 实线表示促进 点线表示抑制 在正常情况下,ACTH对醛固酮的分泌并无调节作用,但切除垂体后,在应激醛固酮的分泌反应减弱,提示在应激情况下,ACTH对醛固酮的分泌可能起到一定的支持作用。 二、肾上腺髓质 肾上腺髓质嗜铬细胞分泌肾上腺素(epinephrine,E)和去甲肾上腺素(norepinephrine,NE)都是儿茶酚胺激素。 (一)髓质激素的合成与代谢 髓质激素的合成与交感神经节后纤维合成去甲肾上腺素的过程基本一致,不同的是在嗜铬细胞胞浆中存在大量的苯乙醇胺氮位甲基移位酶(phenylethanolamine-N-methyltransferase,PNMT),可使去甲肾上腺素甲基化而成肾上腺素。合成髓质激素有原料分为酪氨酸,其合成过程为:酪氨酸→多巴→多巴胺→去甲肾上腺素→肾上腺素,各个步骤分别在特异酶,如酷氨酸羟化酶、多巴脱羟酶、多巴胺β-羟化酶及PNMT的作用下,最后生成肾上腺素。 肾上腺素与去甲肾上腺素一起贮存在髓质细胞的囊泡里内,以待释放。髓质中肾上腺素与去甲肾上腺素的比例大约为4:1,以肾上腺素为主。在血液中去甲肾上腺素除由髓质分泌外,主要来自肾上腺素能神经纤维末梢,而血中肾上腺素主要来自肾上腺髓质。 在体内的肾上腺素与去甲肾上腺素通过单胺氧化酶(monoamine oxidase,MAO)与儿茶酚-O-甲基移位酶(catechol-O-methyltransferase,COMT)的作用而灭活。 (二)髓质激素的生物学作用 髓质与交感神经系统组成交感-肾上腺髓质系统,或称交感-肾上腺系统,所以,髓质激素的作用与交感神经紧密联系,难以分开。生理学家Cannon最早全面研究了交感-肾上腺髓质系统的作用,曾提出应急学说(emergency reaction hypothesis),认为机体遭遇特殊情况时,包括畏惧、剧痛、失血、脱水、乏氧、暴冷暴热以及剧烈运动等,这一系统将立即调动起来,儿茶酚胺(去肾上腺素、肾上腺素)的分泌量大大增加。儿茶酚胺作用于中枢神经系统,提高其兴奋性,使机体处于警觉状态,反应灵敏;呼吸加强加快,肺通气量增加;心跳加快,心缩力增强,心输出量增加。血压升高,血液循环加快,内脏血管收缩,骨骼肌血管舒张同时血流量增多,全身血液重新分配,以利于应急时重要器官得到更多的血液供应;肝糖原分解增加,血糖升高,脂肪分解加强,血中游离脂肪酸增多,葡萄糖与脂肪酸氧化过程增强,以适应在应急情况下对能量的需要。总之,上述一切变化都是在紧急情况下,通过交感-肾上腺髓质系统发生的适应性反应,称之为应急反应。实际上,引起应急反应的各种刺激,也是引起应激反应的刺激,当机体受到应激刺激时,同时引起应急反应与应激反应,两者相辅相成,共同维持机体的适应能力。 (三)髓质激素分泌的调节 1.交感神经 髓质受交感神经胆碱能节前纤维支配,交感神经兴奋时,节前纤维末梢释放乙酰胆碱,作用于髓质嗜铬细胞上的N型受体,引起肾上腺素与去甲肾上腺素的释放。若交感神经兴奋时间较长,则合成儿茶酚胺所需要的酪氨酸羟化酶、多巴胺β-羟化酶以及PNMT的活性均增强,从而促进儿茶酚胺的合成。 2.ACTH与糖皮质激素 动物摘除垂体后,髓质中酪氨酸氢化酶、多巴胺β-羟化酶与PNMT的活性降低,而补充ACTH则能使这种酶的活性恢复,如给予糖皮质激素可使多巴胺β-羟化酶与PNMT活性恢复,而对酪酸羟化酶未见明显影响,提示ACTH促进髓质合成儿茶酚胺的作用,主要通过糖皮质激素,也可能有直接作用。肾上腺皮质的血液经髓质后才流回循环,这一解剖特点有利于糖皮质激素直接进入髓质,调节儿茶酚胺的合成。 3.自身反馈调节去甲肾上腺素或多巴胺在髓质细胞内的量增加到一定数量时,可抑制酪氨酸羟化酶。同样,肾上腺素合成增多时,也能抑制PNMT的作用,当肾上腺素与去甲肾上腺素从细胞内释入血液后,胞浆内含量减少,解除了上述的负反馈抑制,儿茶酚胺的合成随即增加(图11-20)。
图11-20 肾上腺髓质激素生物合成示意图 PNMT:苯乙醇胺氮位甲基移位酶 (责任编辑:泉水) |
第六节 肾上腺(2)
时间:2006-06-19 20:37来源:大众医药网 作者:admin
顶一下
(2)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 上一篇:第七节 胰岛
- 下一篇:第五节 甲状旁腺和甲状腺C细胞
- 发表评论
-
- 最新评论 进入详细评论页>>
