|
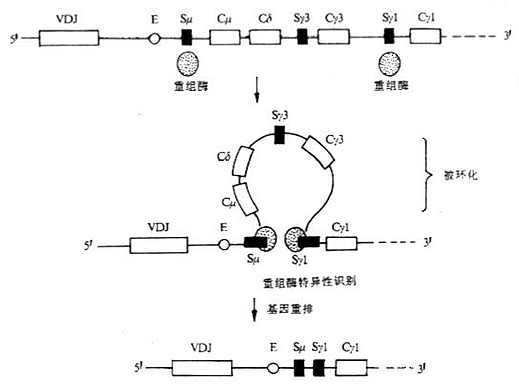
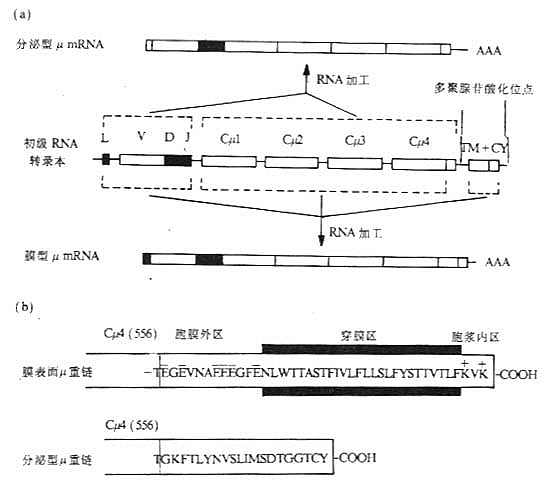
图3-7 Ig类型转换缺失模型 (三)膜表面Ig重链基因 膜表面Ig(mIg)重链基因的外显子结构与分泌性Ig重链的基因外显子结构基本相同,但在基因组的3'端有所不同。作为识别抗原受体的mIg,其重链羧基端有一段疏水性氨基酸插入到胞膜双层脂质中,mIg重链的转录本要比分泌性重链转录本多1~2个外显子,与分泌性重链基因最后一个外显子至少相隔1.4kb,这1~2个外显子编码重链的羧基端部分,这部分氨基酸残基的数目视重链不同而有所差异,如鼠或人mIgμ链的这一部分约为41个氨基酸残基,而鼠mIgε链这部分却有72个氨基酸残基。这个区域可分为三个部分:(1)一个酸性间隔子,靠氨基端侧,与最后一个CH结构域相连,位于细胞膜外侧;(2)跨膜部分,由26个不带电荷的疏水氨基酸形成一个α螺旋,穿过胞膜的脂质双层;(3)胞浆内羧基端部分,3~28个氨基酸残基不等,可能与信号传递有关。
图3-8 RNA的差异拼接与IgM和IgD共同表达在一个B细胞上 分泌形式的μ、α和δ重链最后一个结构域后有一段额外延长的由带电荷氨基酸组成的序列,称为尾片(tail pieces),约含20个氨基酸,在IgM和IgA分子中,单体Ig尾片通过二硫键相互连接或与J链形成二聚体或多聚体。分泌形式μ链的mRNA含有V、D、J、Cμ1、Cμ2、Cμ3、Cμ4和Cμ43'端一个小的外显子转录区。 Ig重链基因除L、V、D、J和C基因片段外,在内含子中还有一些与mRNA转录和免疫球蛋白类的转换有关的结构。 (1)插入顺序(intervening sepuence,IS):有IS1、IS2、IS3……等。 (2)转换区序列:即S区(switch region)。除Cδ外,各个恒定区基因片段上游都有一个同源重复序列的S区,约占2~10kb,分别命名为Sμ、Sγ、Sα和Sε等,与Ig类和亚类的转换有关。S区含有众多串联的高度保守的DNA重复序列,每个重复序列可长到52bp,其确切的功能还不清楚。如Sμ的结构为[(GAGCT)nGGGGT]m,n一般为2~5,但有时可达17,m可达150。目前关于H链转换区的研究主要以小鼠为模型,如4个Sγ是由49bp长的基序的重复序列所组成。Sα至少由长为80pb的15个重复序列,Sε的重复序列长为60bp。在小鼠,B淋巴细胞经T细胞非依赖性活化后常发生Sμ/Sγ3和Sμ/Sγ2b的重组,因此在无T细胞存在条件下,LPS活化小鼠B细胞成为淋巴母细胞,主要转换Ig的亚类为IgG3和IgG2b。 (3)启动子:靠近每个V基因转录位点的上游,含有TATA盒,控制RNA多聚酶Ⅱ作用下的转录过程。大多数Ig的启动子含有许多DNA序列,包括一个保守的8个核苷酸序列,可能是核DNA结合蛋白(nuclear DNA-binding proteins)结合部位,在转录过程中起着重要作用。由于核DNA结合蛋白是由其它基因所编码,因此这种作用方式被称为反式作用(transacting).
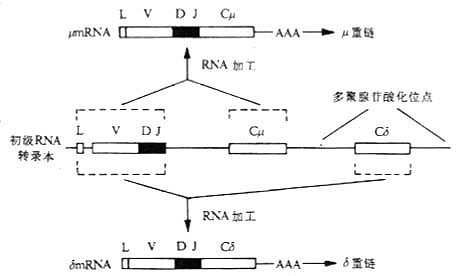
图3-9 膜表面和分泌的Igμ基因及μ链羧基端结构的比较 注:(a)初级RNA转录本通过不同的剪接加工,分别形成分泌型μmRNA和膜型μmRNA。 (4)增强子(E):在人和小鼠和重链和κ轻链中发现有增强子,其核苷酸序列为TGGTAAG。在H链基因中,增强子位于JH3'端侧。当H链VDJ或κ链VJ重排后,V基因上游的启动子靠近增强子,使V(D)JC基因重排后开始更为有效的转录。增强子作用方式与启动子不同,呈方向非依赖方式(orientation-independent manner),即增强子可作用于被转录基因的上游或下游。此外,增强子的作用是细胞特异性。 已重排Ig基因的转录起始于TATA盒下游约20核苷酸处,转录至C基因后,产生初级RNA(primary RNA)转录本,在3'端切断后进行多聚腺苷酸化(polyadenylation),剪接掉不编码的内容含子,如J区与C区之间的不编码序列是在RNA水平上被剪接掉。
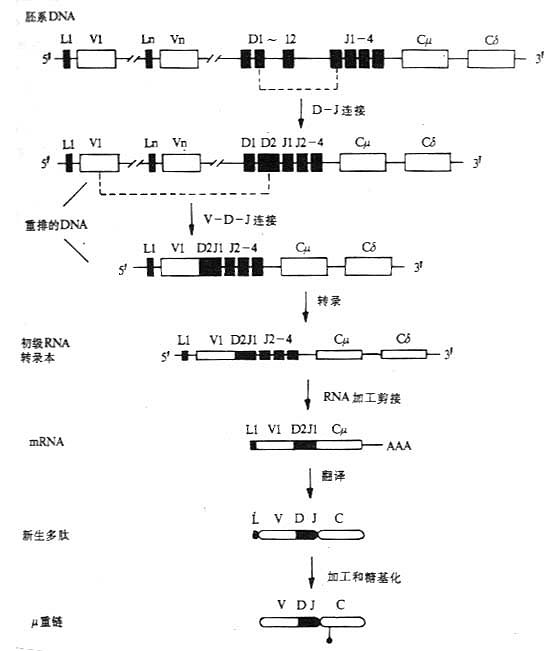
图3-10 小鼠μ链的基因重排顺序、转录和合成 (四)重链基因重排、转录和多肽链的合成。 以μ链基因的重排顺序、转录和μ链合成为例,见图3-10。 (责任编辑:泉水) |
第二节 免疫球蛋白基因的结构和多样性(2)
时间:2006-06-29 00:12来源:大众医药网 作者:admin
顶一下
(4)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 上一篇:二、Ig轻链基因的结构和重排
- 下一篇:三、CD、粘附分子与Ig超家族的关系
- 发表评论
-
- 最新评论 进入详细评论页>>
