|
第三节 膜受体介导的信号转导 与脂溶性的化学信号不同,亲水性信号分子(所有的肽类激素、神经递质和各种细胞因子等)均不能进入细胞。它们的受体位于细胞表面。这些受体与信号分子结合后,可以诱导细胞内发生一系列生物化学变化,从而使细胞的功能如生长、分化及细胞内化学物质的分布等发生改变,以适应微环境的变化和机体整体需要。这一过程可以称之为跨膜信号转导。在这一信号转导过程中,信号分子不进入细胞。虽然有些信号分子与受体结合后可以发生内化(internalization),但这不是主要的作用方式。这种位于膜表面的受体所介导的信号传递主要表现为,各种参与信号传递的信号分子的构象、浓度或分布发生变化,各种信号分子之间发生相互识别和相互作用。 一、膜受体的分类 随着越来越多的膜表面受体被纯化,其结构及转导信号的方式逐步得以阐明。目前,按照受体的结构及其作用方式可将其分为三大类。这三大类受体在配体种类、受体的一般结构和功能及细胞对之发生反应的方式上有所不同,见表21-2。? Table 21-2 Classification of Membrane Receptors:Characteristics of Three Groups of Receptors
二、膜受体信号转导的分子机理 (一)离子通道型受体及其信号转导 离子通道型受体是一类自身为离子通道的受体。这种离子通道与受电位控制的离子通道及受化学修饰调控的离子通道不同,它们的开放或关闭直接受配体的控制,其配体主要为神经递质。
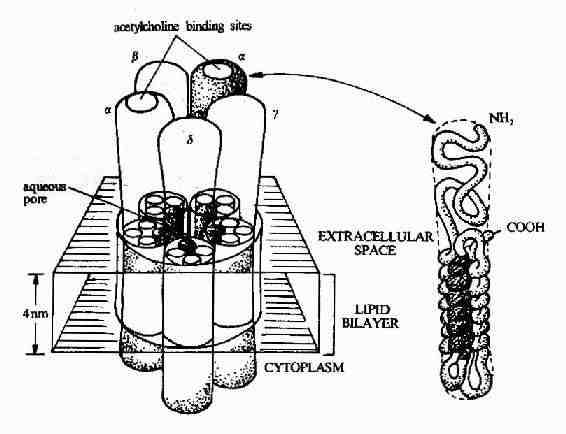
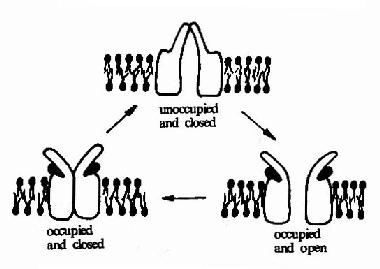
图21-8 乙酰胆碱受体的结构模式图 图21-8显示了作为离子通道受体的典型代表-乙酰胆碱受体的结构模式。乙酰胆碱受体是由5个同源性很高的亚基构成,包括2个α亚基,1个β亚基,1个γ亚基的和1个δ亚基。每一个亚基都是一个四次跨膜蛋白,分子量约60kd,约由500个氨基酸残基构成。推测跨膜部分为四条α螺旋结构,其中一条α螺旋含较多的极性氨基酸,就是由于这个亲水区的存在,使五个亚基共同在膜中形成一个亲水性的通道。乙酰胆碱的结合部位位于α亚基上。? 乙酰胆碱受体可以以三种构象存在(图21-9)。两分子乙酰胆碱的结合可以使之处于通道开放构象,但即使有乙酰胆碱的结合,该受体处于通道开放构象状态的时限仍十分短暂,在几十毫微秒内又回到关闭状态。然后乙酰胆碱与之解离,受体则恢复到初始状态,做好重新接受配体的准备。
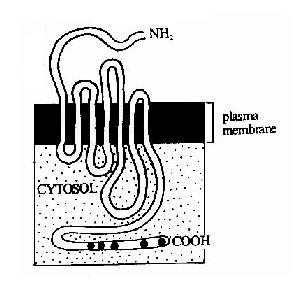
图21-9 乙酰胆碱受体的三种构象示意图 离子通道受体信号转导的最终作用是导致了细胞膜电位的改变,可以认为,离子通道受体是通过将化学信号转变成为电信号而影响细胞的功能的。 离子通道型受体可以是阳离子通道,如乙酰胆碱、谷氨酸和五羟色胺的受体,也可以是阴离子通道,如甘氨酸和γ-氨基丁酸的受体。 (二)G蛋白偶联型受体及其信号转导 G蛋白偶联型受体包括多种神经递质、肽类激素和趋化因子的受体,在味觉、视觉和嗅觉中接受外源理化因素的受体亦属G蛋白偶联型受体。这类受体在结构上均为单体蛋白,氨基末端位于细胞外表面,羧基末端在胞膜内侧。完整的肽链要反复跨膜七次(图21-10),因此亦有人将此类受体称为七次跨膜受体。由于肽链反复跨膜,在膜外侧和膜内侧形成了几个环状结构,它们分别负责与配体(化学、物理信号)的结合和细胞内的信号传递。其胞浆部分可以与一种GTP结合蛋白(简称G蛋白)相互作用,这种G蛋白是该信号传递途径中的第一个信号传递分子,这也是这类受体被称为G蛋白偶联型受体的原因。
图21-10 七次跨膜受体-G蛋白偶联 ?型受体跨膜结构示意图 G蛋白偶联受体的信号传递过程包括(1)配体与受体结合,(2)受体活化G蛋白;(3)G蛋白激活或抑制细胞中的效应分子;(4)效应分子改变细胞内信使的含量与分布,(5)细胞内信使作用于相应的靶分子,从而改变细胞的代谢过程及基因表达等功能。本节将逐一介绍这一过程的主要环节。 1.G蛋白的循环或活化(G Protein Cycle) G蛋白偶联型受体的信号转导途径中的第一个信号传递分子是G蛋白,其活化过程称为G蛋白循环。 G蛋白以α、βγ亚基三聚体的形式存在于细胞质膜内侧。α亚基已发现有20余种,分子量为36~52kd。α亚基具有多个活化位点,其中包括可与受体结合并受其活化调节的部位、与βγ亚基相结合的部位、GDP或GTP结合部位以及与下游效应分子相互作用的部位等等。α亚基还具有GTP酶活性。α亚基结合GDP时是无活性状态,而与GTP结合时则为有活性状态,GTP的水解又使其返回无活性状态。 G蛋白中的β和γ亚基亦有数种,但不及α亚基种类多。在细胞内,β和γ亚基形成紧密结合的二聚体,只有在蛋白变性条件下方可解离,因此可以认为它们是功能上的单体。βγ亚基的主要作用是与α亚基形成复合体并定位于质膜内侧。近年来的研究表明,βγ亚基亦可作用于其下游效应分子。
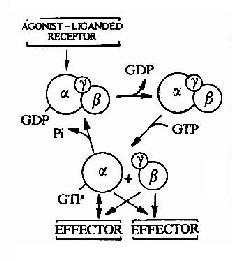
图21-11 G蛋白循环示意图 G蛋白循环的具体过程可见图21-11。当物理或化学信号刺激受体时,受体活化G蛋白使之发生构象改变。α亚基与GDP的亲和力下降,结合的GDP为GTP所取代。α亚基结合了GTP后即与βγ亚基发生解离,成为活化状态的α亚基。活化了的α亚基此时可以作用于下游的各种效应分子。这种活化状态将一直持续到GTP被α亚基自身具有的GTP酶水解为GDP。一旦发生GTP的水解,α亚基又再次与βγ亚基形成复合体,回到静止状态,重新接受新的化学信号。 (责任编辑:泉水) |
第三节 膜受体介导的信号转导
时间:2006-06-20 10:46来源:大众医药网 作者:admin 点击:
384次
顶一下
(3)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 上一篇:第二十二章 癌基因与抑癌基因
- 下一篇:第二节 细胞内受体的信号转导机理
- 发表评论
-
- 最新评论 进入详细评论页>>
