除通过打开突触后膜阴离子(氯)通道外,抑制性神经递质还可以通过关闭突触前膜阳离子(钙)通道、关闭突触后膜阳离子(钙)通道以及打开突触后膜阳离子(钾)通道来启动抑制机制。
γ-氨基丁酸
γ-氨基丁酸是中枢神经系统中主要的抑制性神经递质。虽然它并非人体中常见的二十种氨基酸之一,但由于其结构与谷氨酸的相似性,我们仍称其为一种抑制性氨基酸神经递质。除几种特例,γ-氨基丁酸大部分时间会作用在大脑中的γ-氨基丁酸能神经元(一种小型中间神经元)上来抑制神经回路。
γ-氨基丁酸A型受体
在突触后,γ-氨基丁酸可以作为两种受体的内源性配体,其中我们首先讨论的是γ-氨基丁酸A型受体——一种配体门控阴离子通道。γ-氨基丁酸与其结合后,会增加携带负电荷的氯离子朝向细胞质的涌入,最终导致突触后膜的超极化(抑制性突触后电位)。此受体除了γ-氨基丁酸以外,还拥有许多结合域。类固醇、巴比妥酸盐、苯二氮卓类药物(如阿普唑仑与安定剂)都可以作用在γ-氨基丁酸A型受体上。
γ-氨基丁酸B型受体
γ-氨基丁酸还可以与另一种受体结合——即B型受体。此受体为一种G蛋白耦联受体,也是本文详细讨论的受体之一。γ-氨基丁酸B型受体在神经突触前后都有所表达。当γ-氨基丁酸与神经突触前的B型受体结合后,此受体的活化会诱导与其耦联的G蛋白的Gα_i/o下游通路,并最终导致环磷酸腺苷浓度的下降。突触前轴突终末内环磷酸腺苷浓度的下降会避免突触小泡和突触前膜融合,并同时避免神经递质从突触前膜中分泌出来。当与B型受体耦联的G蛋白被活化时,其α亚基与βγ复合物会解离,释放出的βγ复合物会直接与电压门控钙通道结合并使其闭合,从而抑制钙离子的内涌及其下游神经递质的释放。此外,βγ复合物还会间接地与SNARE蛋白结合,进一步抑制下游神经递质的分泌。
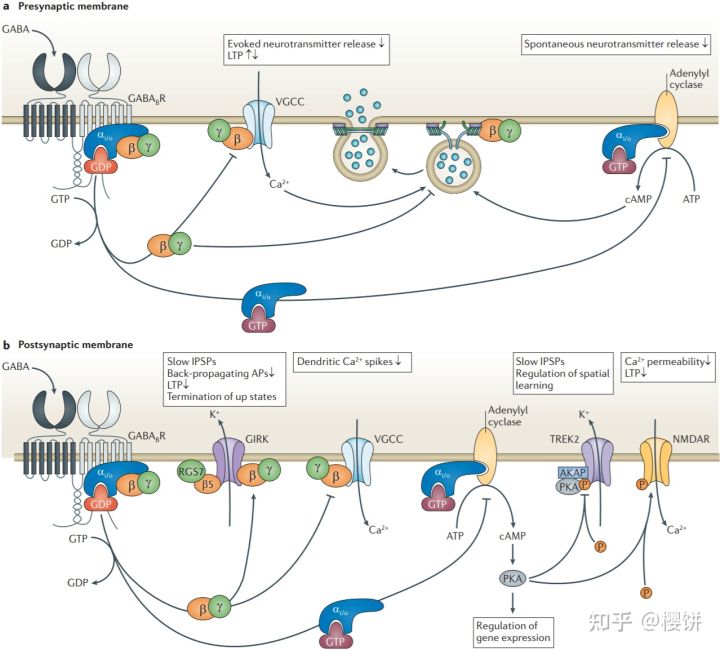
参考
- Rost, B. R. et al. Activation of metabotropic GABA receptors increases the energy barrier for vesicle fusion. J. Cell. Sci. 124, 3066–3073 (2011).
- Sakaba, T. & Neher, E. Direct modulation of synaptic vesicle priming by GABAB receptor activation at a glutamatergic synapse. Nature 424, 775–778 (2003).
- Wells, C. A., Betke, K. M., Lindsley, C. W. & Hamm, H. E. Label-free detection of G protein-SNARE interactions and screening for small molecule modulators. ACS Chem. Neurosci. 3, 69–78 (2012).
- Deng, P. Y. et al. GABAB receptor activation inhibits neuronal excitability and spatial learning in the entorhinal cortex by activating TREK-2 K+ channels. Neuron 63, 230–243 (2009).
- Chalifoux, J. R. & Carter, A. G. GABAB receptors modulate NMDA receptor calcium signals in dendritic spines. Neuron 66, 101–113 (2010).
- Xie, K. et al. Gβ5 recruits R7 RGS proteins to GIRK channels to regulate the timing of neuronal inhibitory signaling. Nature Neurosci. 13, 661–663 (2010).