| Figure 6. [Click image to enlarge] The corollary discharge. Diagrammatic representation of the brain and spinal cord showing the possible levels of corollary discharge by which motor output from the cortex acts on incoming kinesthetic signals arising from the sensory neurones. Corollary discharges are obtained from motor commands and they can influence perception either by modifying incoming sensory signals (in this case at the level of the thalamus) or by acting independently of the incoming sensory signals. Kinesthesia is the sensation by which body weight, position, muscle tension and movement are perceived. Corollary discharges can alter the way in which such kinesthetic signals arising from sensory endings in muscles are interpreted. Such sensory endings in muscle spindles may send signals relating to the length and velocity of movement of a particular set of muscles; these spindles will also send signals arising from their being activated by a certain class of motoneurones in the spinal cord called gamma motoneurones. ]be signals due to the gamma activation of the spindles are removed by a corollary discharge, which at the same time allows the signals from the spindles due to the length and velocity changes to be perceived. In the example shown gamma motoneurones are activated from the motor cortex giving rise to spindle receptor discharges; these discharges together with the additional discharges due to the contraction of the muscles are received by the sensory neurones and transmitted through the group of neurones constituting the gracilis and cuneatus to the thalarnus and thence to the somatosensory cortex; here they give rise to the sense of movement of the muscles. However the initial motor discharge of impulses gates out the sensory discharge relating to gamma motoneurones exciting the spindle receptors; this gating may occur at the many regions of interaction between motor and sensory pathways in the brain and are shown here as occurring in the thalamus for definiteness only. |
In order to make these ideas of Humphreys clear it is necessary to look in detail at the functioning of the human nervous system. First of all what are collateral effects and can they operate in such a way as to modify sensory signals? One of the simplest motor acts that engages the brain and the spinal cord is shown in Figure 6: here a sensory stimulus, such as that arising from sensory spindle receptors in muscle cells concerned with indicating the length and velocity of shortening of the muscle, is relayed through the sensory neurones just outside the spinal cord to the nerve cells just inside the cord within an area called the substantia gelatinosa; these signals are then sent to the main relay station for sensory activity propagating between the spinal cord and the brain, in the group of neurones called the nucleus gracilis and nucleus cuneatus; from there the signals are sent to the thalamus in the brain, which is the receiving area for nearly all the sensory input to the overlying mantle of the brain or cortex; finally the thalamus projects the information to that part of the cortex which is called the somatosensory area, concerned with the analysis of information derived from sensory receptors in the limbs. The kind of information in the signals processed by the somatosensory cortex may require that the muscles that gave rise to the sensory input in the first place be contracted. In this case neurones in the part of the cortex concerned with contracting muscles, namely the motor cortex, project a signal to the appropriate motoneurones in the spinal cord connected to these muscles. These are of two different kinds, namely alpha motoneurones that are attached to cells in the muscle in question that produce the force; the other kind are the gamma motoneurones that are connected with cells that contract in the sensory receptor apparatus itself; contraction of these cells changes the characteristics of the sensory receptors so that they rapidly send signals to the sensory neurones outside the spinal cord and from there to the alpha motoneurones, leading to the contraction of the bulk of the muscle cells; they also send signals to the sornatosensory cortex via the thalamus along the pathway already described. Two kinds of information are then sent via the sensory neurones to the brain: one of these relates to the signal arising from the sensory receptors in the muscle concerned with the position, tension and movement of the muscle, known collectively as kinesthesia; the other relates to the signal arising from the receptors as a consequence of their being contracted by the gamma motoneurones. This latter signal is gated out before it reaches consciousness by a collateral signal from the motor pathway as shown in Figure 6; the sensory signal concerned with the state of kinesthesia of the muscle is not gated out, but is allowed to reach consciousness. The level in the brain or spinal cord at which this gating procedure is carried out is not known; it is shown to occur at the level of the thalamus in Figure 6 simply for the sake of definiteness. Humphrey is therefore correct in his assertion that modification of sensory signals can occur before they reach consciousness as a consequence of a collateral effect from the motor pathway.
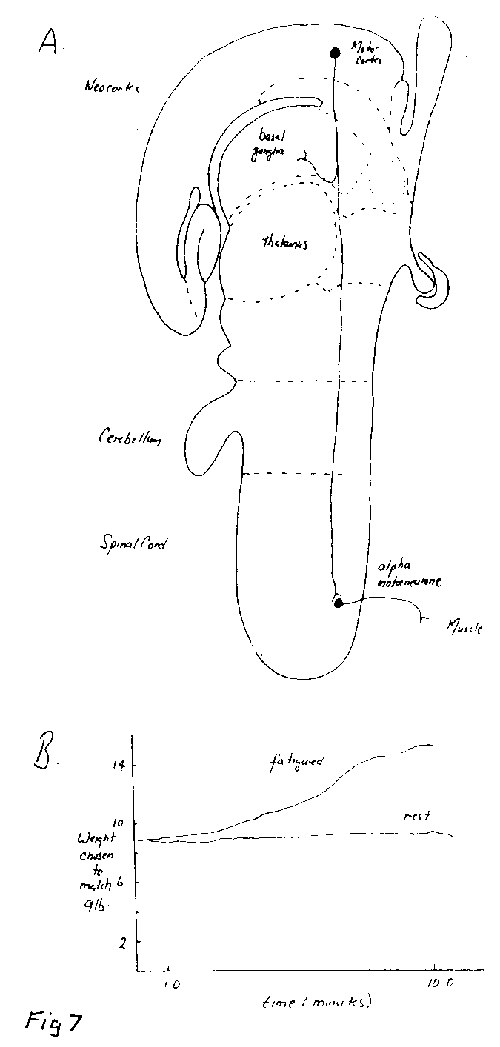 |
Figure 7. [Click image to enlarge] The corollary discharge. This figure gives a diagrammatic representation of the brain and spinal cord illustrating a possible output from the motor cortex responsible for the perception of heaviness of a held object. For definiteness this corollary discharge is shown at the level of the basal ganglia. Such discharges can give rise to the sensation of muscular effort as occurs when lifting and supporting an object. In the example shown neurones in the motor cortex (called Betz cells) that project to the motoneurones in the spinal cord are illustrated; Betz cells may be activated to contract muscles involved in lifting the limbs or an object such as a suitcase; when they do this a corollary discharge is sent (at the level of the basal ganglia?) which gives rise to the perception of heaviness of the limb or suitcase, and this is simply related to the extent of the motor discharge that occurs. Subjects that experience a stroke may have to send a larger than normal discharge down the remaining functional Betz cells to achieve the aim of lifting their arms and so experience them as as an enormous burden. The graph shows the results of an experiment in which the subject has to support a 9 lb. weight with one arm ( the experimental arm) while being asked at intervals to choose what they thought were equal weights to be supported in the same way by the other arm (the control arm). When the experimental arm was allowed to rest between the trials the subject choose weights with the control arm close to the 9 lb weight held by the experimental arm (see 'rest curve'). If however the experimental arm had to support the 9 lb weight continuously, then the subject choose weights with the control arm that were successively greater (see 'fatigued ' curve) than the 9 lb weight indicating the increased sense of heaviness. This arises from the increase in corollary discharge with time as muscles have to receive a greater discharge to support the weight continuously. The perception of heaviness does not arise from sensory signals in the muscle being relayed back to the brain. This graph is due to experimental work of McCloskey, Ebeling and Goodwin carried out in 1974. |
Corollary discharges from the motor pathway can also be used to generate a sensation independent of any incoming sensory signals. They can generate sensations of muscular force or heaviness although they cannot generate sensations of movement. Figure 7A shows how the sensation of the heaviness of an object held in the hand is generated by a collateral effect. The pathway from the motor cortex to the alpha motoneurones is shown to give off a collateral branch at the level of the basal ganglia; this, it is hypothesized, can generate a sensation in the cortex of the degree of heaiviness of the object by firing impulses in proportion to those that are being propagated down the motor pathway. It follows that when a muscle is weakened by fatigue, such as when holding a heavy suitcase, a greater number of impulses are required by the non-fatigued component of the muscle in order for the muscle to continue lifting the suitcase; the collateral then receives a greater number of impulses and so a greater sensation of heaviness is experienced. An experimental example of this is given in Figure 7B[;] here a comparison is made between the extent to which a subject perceives the heaviness of a suitcase held continually in one hand by comparing it with a known weight held in the other for a short period of time. The graphs show that in this matching experiment the known weight chosen to be equivalent to the suitcase gradually increases over time, indicating the increased sensation of heaviness. This sensation is due to the collateral effect.
Humphrey is correct then in his suggestion that collateral effects can modify both the kinds of sensations that enter consciousness as well as generate sensations that did not arise from the workings of our sensory receptors. Figure 7 summarizes the situation. Humphrey speculates that early during evolution nerve pathways were layed [sic] down that allowed an animal to respond to say a noxious stimulus to the skin by 'wriggleing' away; in higher vertebrates this simplest pathway might consist of the primary sensory neurones just outside the spinal cord that receive information concerning noxious stimulation projecting to upper motoneurones in the reticular formation wbich then project down to the lower motoneurones and from there to the muscles which are to be contracted to produce "wriggleing" (Figure 8A). At a later stage of evolution mechanisms were put in place that allowed the nervous system to 'gate' out sensory information, using projections from the brain to the sensory gate-way to the cortex , the thalamus, as shown in Figures 8B and 8C. We have already seen how information gathered by primary sensory neurones concerned with muscle receptors can be gated out before it reaches the somatosensory cortex by means of a collateral feedback from the motor cortex at the level of the thalamus (Figure 8B). Such a feedback could occur via the well known pathway from motor cortex to basal ganglia and from there to the reticular nucleus that lies just outside the thalamus; this then projects to the somatosensory cortex (Figure 8B). Sensory information that is gathered by the retina is also 1 gated' as it passes through the thaIamus on the way to the visual cortex, as shown in Figure 8C. The primary visual pathway is from the retina to the thalamus and from there to the visual cortex; neurones exist in the cortex that project back to the thalamus where they can gate the incoming visual information (Figure 8C). There is then evidence for both the modulation of signals arising from the primary sensory neurones as well as from the visual sensory neurones at the level of the thalamus. In this way the brain can determine the sensory information which reaches it.
(责任编辑:泉水)