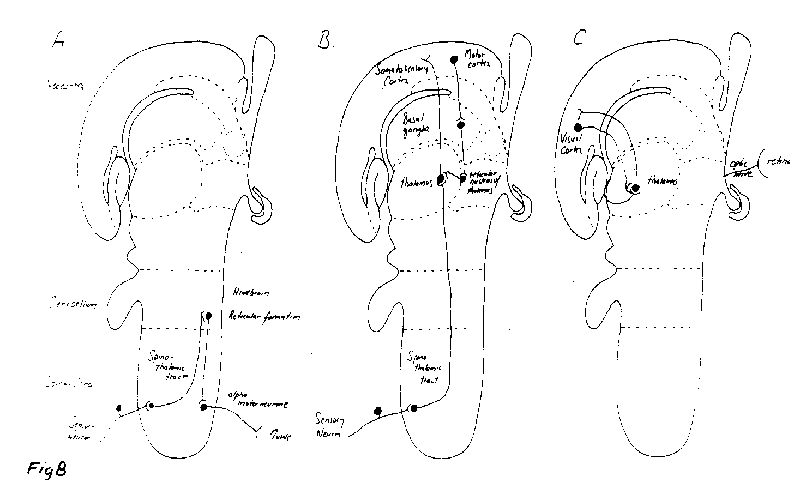
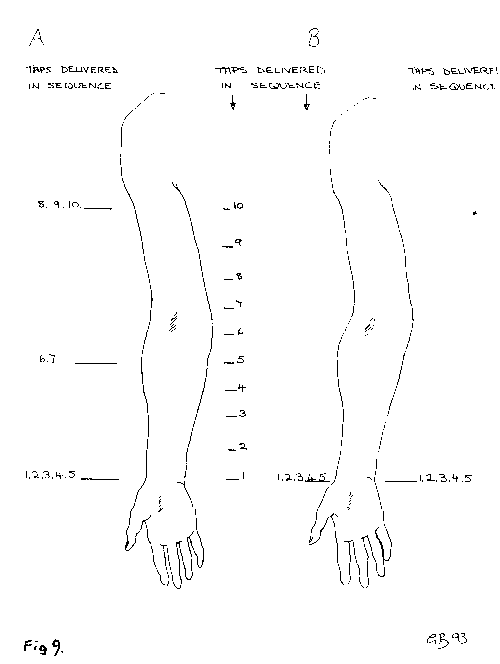
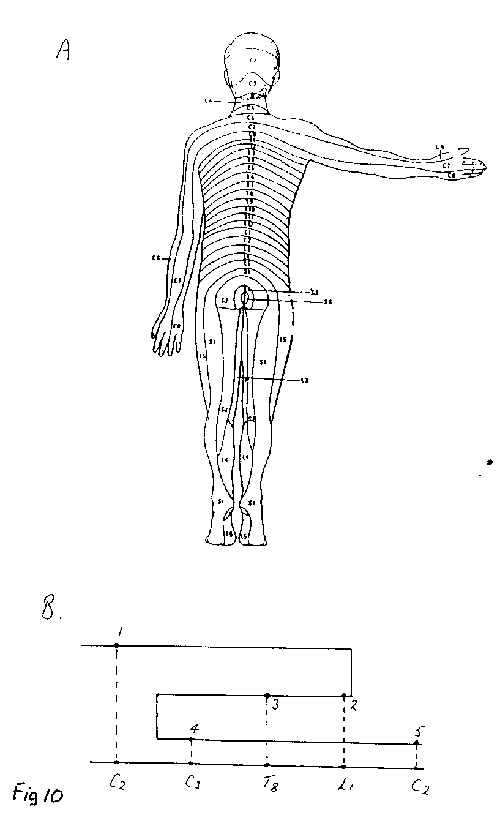
[NextPage] The question arises as to whether or not the modulatory effect of collaterals on the sensory information passing through the thalamus simply involves just a gating operation, that is the removal of information. We have already seen that this is not the case as a collateral effect generates the experience of heaviness when holding a suitcase, independent of any incoming sensory impulses (Figure 7). Humphrey suggests that collaterals may also sustain impulse traffic in sensory nerves after the sensory perception has passed. The effect of this would be to experience sensations without there be any continuing effect on sensory receptors, although these receptors would have been involved in the initiation of the experience in the first place. This brings up a question concerning the time over which consciousness of a sensation occurs. The tricky nature of the experience of time in consciousness as compared with objective time, measured by a clock for instance, is well illustrated by the 'cutaneous rabbit' perceptual illusion (Figure 8). In this illusion a series of taps to the wrist is followed by taps to the upper forearm and then to the shoulder, as shown in Figure 9A. Surprisingly this is experienced as a series of taps that are equally spread out along the whole length of the arm, rather than confined to just three positions on the arm, as if an animal (a 'rabbit') had run up the arm. Even more surprising is the result of just giving the series of taps to the wrist in the absence of any taps to the forearm or the shoulder (Figure 9B): in this case the taps are all experienced as confined to the wrist without any of them appearing to be spread out along the arm. Why then in the first experiment did the brain interpret the taps at the wrist as experienced spread out along the arm whereas in the second case they remain confined at the wrist? With reference to Figure 8, a plausible explanation why the first five taps in A were experienced as distributed along the arm whereas the five taps in B were confined to the wrist is that the taps are not perceived simultaneous with the events. In a certain window of time (1 to 2 seconds) the brain determines the most likely spacetime story relating to the taps: in A the preliminary story that all five taps occur at the wrist is wiped out by the later arriving taps so that the final story that enters consciousness is that the taps are spread out equally in a space-time sequence; in B the preliminary story that all five taps occur at the wrist is not wiped out by any later events and so this enters consciousness. The brain then uses the time available before behavour is acted out to arrive at the most reasonable story based on sensations (the taps) and past experience to arrive at an interpretation. This window in time could be delineated by the earliest time at which sensations enter the brain and the latest time at which the experiences might be used to modify behavour. The actual time at which occurrences are first registered in the brain might not then be the same as the times allocated to them by consciousness. Another example of this is illustrated in Figure 10A, which shows the distribution of dermatomes for skin sensations as in Figure 4. The nerves leading from the dermatomes over the buttocks to the brain clearly involve a much longer pathway than do the nerves from the dermatomes over the neck to the brain. It might be naively expected then that if one was to be touched simultaneously on the buttocks and the neck, according to objective timing, then the experience of being touched on the neck would enter consciousness before that of being touched on the buttocks. But this is not the case, as it depends on the context in which this touching occurs as to whether one has the conscious experience of being touched in one place or the other within a certain window of time The hypothetical graph in Figure 10B illustrates that the time of experiencing being touched on various parts of the body (or on different dermatomes) need not coincide with the objective time of the sequence of touchings. The brain creates the most likely story, using the information that it receives from sensory receptors, the context in which this is gathered, and past experience, before allocating times to particular events. Humphrey suggests that collaterals not only gate incoming sensory activity, for example at the level of the thalamus, but they can also sustain that activity after the sensory receptors are no longer stimulated. This would then give rise to a sensation that is extended in time within consciousness. It gives rise to an important idea in Humphreys' scheme, namely that of the 'sustained sentient loop', in which the issuing of an outgoing command over a collateral can give rise to a sensation that is extended over time in consciousness by the sustained activity of the collateral.
顶一下
(19)
100%
踩一下
(0)
0%
------分隔线----------------------------
|