|
基底神经环路(七)——基底神经环路对一个运动动作的控制过程。 摘要:本篇以基底神经环路用于对一组主动肌和拮抗肌进行信号控制的两条神经通道为例,从神经元的层面描绘出其局部神经投射环路,并以此描述基底神经环路对一个运动动作的具体控制过程。 ———————————————— 7.1机体在同一关节两个相反方向上的运动,需要由至少两条作用力方向相反的骨骼肌进行控制,也即关联的主动肌和拮抗肌,而在基底神经环路,存在关联的一组神经通道对这两条骨骼肌进行控制。根据前面几篇文章所分别描述的纹体(新纹状体)、苍白球、底丘脑、黑质等各个核团的构成、工作机制及彼此的神经投射关系,描绘出基底神经环路用于对一组主动肌和拮抗肌进行信号控制的一组神经通道的神经元投射关系,如图7—1。本篇以此来详细描述基底神经环路对运动动作的具体控制过程。 需要说明的是:⑴、这些神经通道并非是某几个神经元固定对应于某条骨骼肌,而是可以根据运动的相关性,整合相关的神经元形成一条神经通道来控制一条骨骼肌。⑵、从纹体到苍白球是多数神经元向少数神经元的聚合投射,不同时刻可由不同的神经元参与信息整合,而且,中枢神经也需要具备一定的冗余度和代偿性,所以,通道中的各个神经元节点可能是由多个神经元共同参与工作的,但为了更简明来说明工作原理,图7—1仍以单个神经元进行投射来描述。⑶、中枢神经元的活动状态,除了其自身存在低节律的没有信号输出价值的自发放现象,其是否进行高节律的激活发放并进行有价值的输出,取决于其接受输入的兴奋性信号和抑制性信号的整合。⑷、所谓的低节律或高节律发放都是相对自己的,不同核团和不同神经元在工作时其发放节律不一样,其发放节律的绝对数值不能作为比较。⑸、底丘脑和被盖网状核里面的神经结构都不止一级神经元那么简单,但鉴于它们整体作为一个发放兴奋性激励信号的核团,其内部的神经投射对描述外部大环路的工作机制影响较小,所以为了简明也只把它们画成一级神经元。⑹、纹体向苍白球和黑质的GABA神经投射,除了包含GABA递质,还包含有多种神经肽(P物质、强啡肽、脑啡肽等),这些神经肽的具体作用不明,但似乎能够加强纹体GABA神经激活发放时对苍白球和黑质的抑制作用。⑺、本文未考虑来自杏仁核和中缝背核等与情绪和机体状态相关的各种调制性投射。 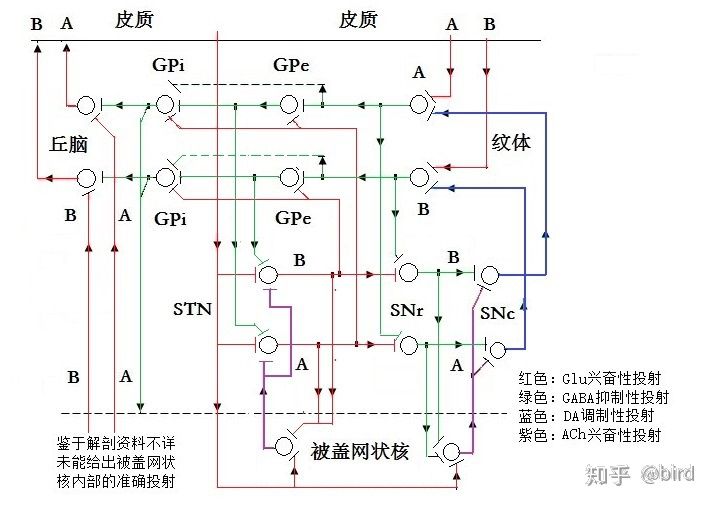 7.2在基底神经环路,两条关联的神经通道A通道和B通道对一组关联的主动肌和拮抗肌的运动动作进行控制的过程,详细描述如下:(由于是从神经元的层面进行细节性的描述,所以会显得比较繁琐)。 ⑴、在静止状态下,纹体上A、B两个通道的输入端都没有得到来自皮质谷氨酸神经的兴奋性运动信号的输入,又都受到来自黑质致密部的多巴胺神经的一定程度的抑制,(纹体还存在兴奋性的胆碱能神经,但这些胆碱能神经属于只在纹体内部进行局部投射的中间神经元,起到内部的信号连接作用),所以,两个通道在纹体的GABA神经元都处于低节律的自发放状态,其输出对苍白球外侧份和黑质网状部只产生弱抑制作用。苍白球外侧份由于受弱抑制而处于高节律发放输出,一方面对底丘脑进行强抑制,使底丘脑—被盖网状核环路的发放节律降低,通过被盖网状核使脑干网状结构相关神经的活动降低,保持静止状态,另一方面对苍白球内侧份进行强抑制,使内侧份神经处于低节律发放,松开对丘脑—皮质上行通道的抑制,也即在静止状态下对上行通道是放行的,所以不会对运动的启动造成影响。 同时,两个通道的纹体GABA神经对黑质网状部的投射都只有弱抑制,使黑质网状部的GABA神经处于较强的活动,其输出对黑质致密部SNc的多巴胺神经产生较强的抑制;黑质致密部SNc的多巴胺神经因此而处于较低强度的活动,对纹体的A通道和B通道存在一定程度的弱抑制作用。这种弱抑制作用既能够使两个通道的纹体GABA神经在没有接收到皮质兴奋信号时,保持在低节律的自发放状态,没有有效输出;又能够使纹体某一通道在接受到皮质兴奋信号时,不会受到过多抑制,所以能够激活进行高节律的发放输出。(如果黑质致密部受损,没有在静止状态下对纹体保持足够程度的弱抑制,则纹体会由于自发放节律过高而产生有效的信号输出,并通过一系列的兴奋和反馈而使两条通道的神经活动产生周期性的振荡现象,导致机体静止性震颤,下一篇文章另有详细描述)。 总之,在静止状态下,基底神经环路A和B两个通道的神经活动状态都差不多,由于皮质没有运动输出,纹体便没有兴奋信号输入,基底环路也没有形成运动控制,两个通道所控制的骨骼肌都维持在一种静止又放松的状态。(这里说的是运动皮质,而不包括小脑,实际上肌肉还受到小脑控制,比如人体站立而静止时,运动皮质没有运动信号输出但小脑有信号输出)。 ⑵、当某一通道比如A通道接收到来自皮质的一串运动信号时,纹体上A通道的GABA神经受到强的兴奋激励,激活输出脉冲串对A通道的苍白球外侧份进行抑制。在脉冲发放期间,苍白球外侧份的GABA神经受到强抑制而活动减弱(降低发放节律),其输出放松对A通道的底丘脑和苍白球内侧份的抑制。底丘脑所受的抑制减弱,其与被盖网状核构成的激励脉冲环路发放节律升高,一方面通过被盖网状核兴奋了运动神经系统其他核团,促使脑干网状结构—丘脑—皮质上行运动信号继续发放,另一方面投射到苍白球内侧份;苍白球内侧份的GABA神经受到底丘脑核高节律的兴奋激励,同时所受外侧份的抑制减弱,于是出现高节律发放,其输出对丘脑→皮质上行投射的A通道进行抑制。而在脉冲的间隔期,苍白球和底丘脑的活动跟静止状态差不多,苍白球内侧份放松对丘脑—皮质上行通道的抑制。这一过程不是单一的动作,也不是一种持续状态,而是动态的节奏性的循环进行:皮质的不同神经元先后激活依次向纹体输出运动信号形成间隔性的脉冲串,纹体也间隔性的向苍白球外侧份输出脉冲串,对苍白球和底丘脑构成的环路的神经活动进行调制。由于纹体输出的脉冲频率较低(1—10赫兹),而苍白球内侧份的放电脉冲频率很高(60—100赫兹,甚至大于100赫兹),最终苍白球内侧份输出的是以内侧份放电为基础、受纹体输出脉冲调制的间隔性的簇状信号,(放电频率60赫兹以上,调制频率1—10赫兹)。这一簇状信号输出到丘脑腹外侧核和腹前核(涉及无意识动作)和板内核(涉及有意识动作)等核团,对丘脑—皮质上行A通道进行间隔性的放行和制动控制,不断上行激励皮质上跟A通道相关的神经活动,使A通道的运动信号持续输出,受该通道控制的骨骼肌持续收缩作为主动肌主导运动动作的进行。 同时,纹体上A通道的GABA神经输出脉冲还对A通道的黑质网状部的神经元进行强抑制,黑质网状部的GABA神经受强抑制而低节律发放,对被盖网状核和黑质致密部的抑制减弱,于是黑质致密部的多巴胺神经激活产生高节律发放,其输出对纹体进行强抑制。而根据前面文章对黑质的分析,纹体与黑质之间的往返投射存在位置上交叉倒置的投射关系,所以,当纹体A通道的GABA神经激活输出对黑质网状部进行强抑制,则导致A通道黑质致密部的多巴胺神经输出反过来是对纹体B通道进行强抑制,使得纹体B通道的GABA神经在A通道激活输出期间受强抑制而没有输出,(更低节律的发放)。于是B通道的苍白球外侧份的GABA神经继续高节律发放,其输出对B通道的底丘脑和苍白球内侧份进行强抑制,使该通道的神经活动减弱,包括由于抑制被盖网状核而抑制了该通道的其他神经核团的活动,最终是使B通道处于静止状态而没有运动输出,受该通道控制的骨骼肌放松,不对A通道控制的主动肌造成对抗。 ⑶、反过来,当皮质向纹体B通道的神经元输出运动信号时,通过同样的控制机制,(只是A和B通道相反),其结果是使B通道的神经活动增强,苍白球内侧份对丘脑→皮质的上行B通道进行间隔性的放行和制动控制,使皮质上与B通道相关的运动神经不断得到兴奋激励而不断进行运动输出,受B通道所控制的骨骼肌作为主动肌持续收缩主导运动动作的进行。同时通过纹体—黑质的交叉倒置投射对A通道的纹体神经进行强抑制,使整个A通道的神经活动减弱,包括由于抑制被盖网状核而抑制了该通道的其他神经核团的活动,最终是使A通道处于静止状态而没有运动输出,受该通道控制的骨骼肌放松,不对B通道控制的主动肌造成对抗。 ⑷、在以A通道控制的肌肉收缩的这一动作的执行过程中,如果感觉皮质没有产生运动到达目的位置的感觉信号,联合皮质的思维系统也没有产生有意识的停止或改变动作的输入信号,则A通道和B通道维持上述的神经活动状态,使得A通道控制的主动肌继续保持收缩状态,B通道控制的拮抗肌也继续保持放松状态,于是动作继续进行。 ⑸、在第⑵点所述的运动动作接近末端或中途需要减速时,联合皮质的思维系统意识到这一点,或者有意识地要停止该动作,或者感觉皮质由于视觉或触觉传入而感觉到运动接近目标位置,或者有来自肌肉或肌梭传感器的到达位置的感觉传入,这些信号会传导到脑干网状结构,引起脑干网状结构(包括被盖网状核)的神经活动产生变化。这种变化可能通过两方面对运动进行影响:一是使原主动肌通道(A通道)底丘脑的激励脉冲发放节律升高,从而使放行段的执行时间减少,使主动肌的运动幅度和速度降低;二是兴奋了拮抗肌的B通道,使B通道的脑干网状结构—丘脑—皮质上行通道出现激励脉冲,激励兴奋了皮质跟拮抗肌相关的神经活动,于是皮质开始向拮抗肌输出运动信号,拮抗肌收缩产生反作用力而使运动减速或刹车。拮抗肌一旦产生运动动作,则同样会通过基底环路进行过程控制。 ⑹、根据运动动作的制动刹车或减速的不同情况,主动肌和拮抗肌的动作变化也会有不同。在粗放的动作需要急剧停止时,由于传入信号变化程度大,基底环路的主动肌和拮抗肌控制通道会进行更快速和更完全的状态切换,使主动肌和拮抗肌快速完成切换以进行快速制动刹车。而在平常的运动末端需要准确定位、或运动中途需要调整运动方向因而需要进行缓慢减速时,如果也这样对两个通道进行简单直接的状态切换,可以想象,这要不就会由于刹车不足而造成动作过头,要不会由于刹车过大而造成动作不足甚至产生动作回弹,这样都会使动作生硬粗糙也容易导致肌肉疲劳,所以,笔者推测在这种状态下,基底环路对两个控制通道进行频繁的高节奏的多次状态切换,使主动肌和拮抗肌进行快速的频繁的交替收缩和放松,产生类似频繁的点刹车式的制动,使得运动动作平稳连贯而且定位准确。而这一使两个通道进行快速的多次状态切换的功能,便需要纹体—黑质环路(可能还有苍白球—黑质环路)的互抑作用来协调完成。这也是黑质受损的帕金森病人,在简单动作的执行过程还比较顺利,而在动作末端或在进行复杂动作,需要两个通道进行频繁切换状态时,便难以顺利进行的原因。(见后面文章关于帕金森的详细叙述)。 按照这样的分析,在运动快速顺畅进行时,基底环路的发放节律反而会比较低,而在运动末端制动或中途减速时,基底环路的整体发放节律将会明显升高,这可以通过实验来观察验证。 7.3总之,基底神经环路对运动的控制,既不是静态的黑盒子理论的易化运动和抑制运动这么简单,也不是线性的准确的步骤控制,而是一种非线性的动态的过程控制。皮质对运动动作的启动是由脑干网状结构←→丘脑←→皮质的系统决定的,但动作一旦启动,基底环路便同时介入动作的过程控制来使动作更平滑和准确。(实际上笔者倾向于认为运动的启动,是由脑干网状结构—丘脑—皮质和被盖网状核—底丘脑—基底环路两方面同时启动的)。如果没有基底环路,借助记忆在皮质上的运动信息也能够进行某些运动,但运动将是粗糙和生硬的。由于神经活动的非线性,基底环路不同核团的神经放电节律的差异也很大,所以基底环路并非对运动动作进行准确的步骤控制,而是通过多个环路之间互相调制彼此的工作状态。当皮质向纹体A通道发放运动信号时,纹体相关神经元被激活进行高节律发放输出,但这并非简单的对运动进行易化或抑制,而是一方面抑制该通道的苍白球外侧份,最终是使底丘脑—被盖网状核环路的神经活动发放节律升高,既通过被盖网状核激励网状结构→丘脑→皮质的上行兴奋信号,又同时使苍白球内侧份产生簇状的脉冲信号去控制这一上行兴奋信号,从而使该通道在交替放行和制动中上行激励皮质,激励该通道相关的运动皮质在受控状态下进行运动输出,驱动主动肌收缩产生运动动作;另一方面是投射到黑质,通过纹体—黑质交叉投射环路去抑制相对的拮抗肌B通道的神经活动,最终是使拮抗肌通道的运动皮质神经活动降低,处于没有运动输出的状态,以配合主动肌的动作顺利进行。 当然,上面是以一组主动肌和拮抗肌为例进行描述,实际上机体的运动往往都需要多组骨骼肌通过协调来完成,所以,基底神经环路实际上是多条神经通道同时在并列工作,不同通道之间也会相互配合。比如,当机体进行一个复杂动作时,实际上是若干骨骼肌作为主动肌(协同肌)在一起收缩,而同时也有若干骨骼肌作为拮抗肌来配合工作。而且,黑质中不同的多巴胺神经对纹体不同GABA神经的抑制作用,以及底丘脑核中不同谷氨酸神经对苍白球中不同GABA神经的兴奋作用,都会由于发放节律的高低而强弱不同,所以,在同一时刻,基底环路的输出对机体各骨骼肌放行和刹车的控制作用都可以是强弱不同的,以此对机体运动的过程进行精细控制,使运动动作平滑连贯而且定位精确。 7.4另外,上面所述的⑴至⑸的控制过程,是按照运动信息的主要的整合通道是从纹体向苍白球外侧份投射,再由苍白球外侧份向内侧份投射来分析的,并认为纹体向苍白球内侧份的直接投射并非运动信息的主要整合通道,更主要是属于神经肽等调制性投射。(详见基底神经环路(四)——苍白球及基底环路对动作幅度的控制的分析)。而如果实际情况不是这样,运动信息的主要整合通道还是大多教材所描述的直接通道,从纹体直接向苍白球内侧份进行投射,则其神经投射关系修改为图7—2,(注意图7—1与图7—2的差别)。变成运动信息由纹体直接向苍白球内侧份进行投射,中间减少一次抑制过程,这会导致内侧份对丘脑—皮质上行通道的放行和刹车的时序(时间点)有变化。但是,对于整个基底神经环路的工作,纹体—苍白球—丘脑这一运动信息整合的主环路及其作用不变,纹体输出对底丘脑—被盖网状核激励脉冲环路的调制及该环路的作用不变,纹体—黑质交叉投射环路及其互抑调制的作用不变,所以基底环路对运动的过程控制和控制结果仍然是一样的。 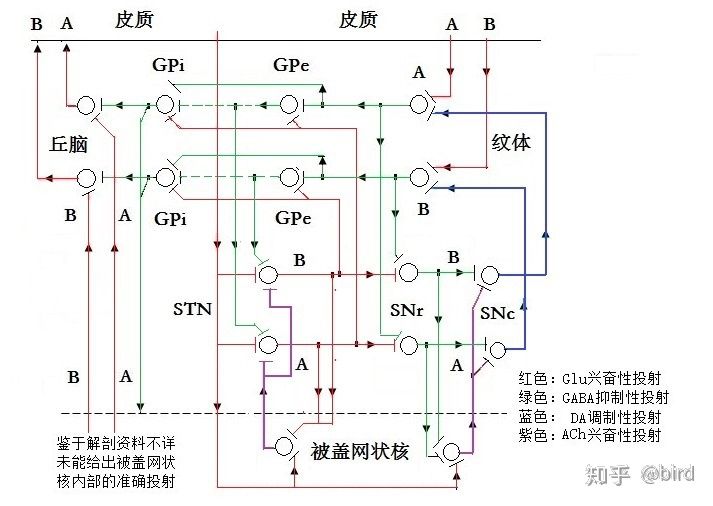 7.5基底神经环路的运动信息整合通道(主环路)是由兴奋性和抑制性氨基酸神经来构成,但黑质却采用多巴胺神经而不是氨基酸神经来实现这种互抑作用,笔者分析其原因可能是调制作用时效性不同的需要。抑制性氨基酸神经虽然也对所投射的神经元起到抑制作用,但抑制作用时效很短,如果用于对两条运动信息整合通道的氨基酸神经进行互相抑制,由于抑制性脉冲与兴奋性脉冲的时效性接近,则很难保证能够对兴奋性信号进行可靠的抑制。而多巴胺递质的抑制作用时效性较长,多巴胺神经脉冲输出所产生的抑制作用时间足够长,(由于缺乏数据,笔者只能大概估计其时间在几十毫秒以上甚至上百毫秒),能够可靠地对一组(几个到几十个,甚至更多)兴奋性脉冲的兴奋作用进行有效抑制,所以能够对两条运动信息整合通道的氨基酸神经进行可靠的互相抑制。如果黑质未能有效进行这种互相抑制,则会出现主动肌和拮抗肌同时收缩互相对抗的异常情况,导致出现肌张力异常的运动障碍,比如帕金森病症,见下一篇文章关于基底节环路和运动障碍的描述。 ——ST量子写于2017年11月。 ———————————————— 1、本文不是资料,而是笔者对基底环路工作机制的分析和猜想。 2、本文原创,但不清楚近期是否有其他文献公开类似内容。 3、欢迎批评指正,或共同探讨。转载请联系作者。 (责任编辑:glia) |
人脑工作机制(27):基底神经环路(七)——“
时间:2022-08-26 11:18来源:网络 作者:叮咚
顶一下
(0)
0%
踩一下
(0)
0%
------分隔线----------------------------
- 发表评论
-
- 最新评论 进入详细评论页>>

- 特别推荐
-

- 推荐内容
-
- 小胶质细胞在人类大脑发育中的关
研究人员通过将小胶质细胞(大脑免疫细胞)整合到实验室培养...
- 呼吸影响瞳孔大小
摘要 研究人员发现,呼吸会影响瞳孔大小,吸气时瞳孔最小,呼...
- 为什么有些人能生动回忆梦境,而
摘要 一项新研究探讨了为什么有些人能生动回忆梦境,而有些人...
- Science Advance:光催化激活神经Ca2+信号
近日,杭师大材化学院刘俊秋教授团队/生科院沈万华教授团队携...
- 体生长抑素信号在大脑中的作用
作者:宾夕法尼亚州立大学 Sam Sholtis 新研究表明,由大脑中许多...
- 协和发现国内首例“阴性艾滋病患
HIV抗体检测为常见的艾滋病筛查手段,阳性一般意味着感染,阴...
- 小胶质细胞在人类大脑发育中的关