|
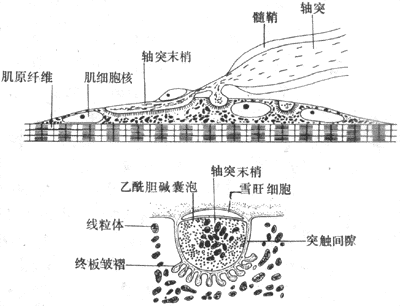
第四节 肌细胞的收缩功能 人体各种形式的运动,主要是靠一些肌细胞的收缩活动来完成的。例如,躯体的各种运动和呼吸动作由骨骼肌的收缩来完成;心脏的射血活动由心肌的收缩来完成;一些中空器官如胃肠、膀胱、子宫、血管等器官的运动,则由平滑肌的收缩来完成。不同肌肉组织在功能和结构上各有特点,但从分子水平来看,各种收缩活动都与细胞内所含的收缩蛋白质,主要与肌凝蛋白和肌纤蛋白的相互作用有关;收缩和舒张过程的控制,也有某些相似之处。本节以研究最充分的骨骼肌为重点,说明肌细胞的收缩机制。 骨骼肌是体内最多的组织,约占体重的40%。在骨和关节的配合下,通过骨骼肌的收缩和舒张,完成人和高等动物的各种躯体运动。骨骼肌由大量成束的肌纤维组成,每条肌纤维就是一个肌细胞。成人肌纤维呈细长圆柱形,直径约60μm,长可达数毫米乃至数十厘米。在大多数肌肉中,肌束和肌纤维都呈平行排列,它们两端都和由结缔组织构成的腱相融合,后者附着在骨上,通常四肢的骨骼肌在附着点之间至少要跨过一个关节,通过肌肉的收缩和舒张,就可能引起肢体的屈曲和伸直。我们的生产劳动、各种体力活动等,都是许多骨骼肌相互配合的活动的结果。每个骨骼肌纤维都是一个独立的功能和结构单位,它们至少接受一个运动神经末梢的支配,并且在体骨骼肌纤维只有在支配它们的神经纤维有神经冲动传来时,才能进行收缩。因此,人体所有的骨骼肌活动,是在中枢神经系统的控制下完成的。 一、神经-骨骼肌接头处的兴奋传递 运动神经纤维在到达神经末梢处时先失去髓鞘,以裸露的轴突末梢嵌入到肌细胞膜上称作终板的膜凹陷中,但轴突末梢的膜和终板膜并不直接接触,而是被充满了细胞外液的接头间隙隔开,其中尚含有成分不明的基质;有时神经末梢下方的终板膜还有规则地再向细胞内凹入,形成许多皱褶,其意义可能在于增加接头后膜的面积,使它可以容纳较多数目的蛋白质分子,它们最初被称为N-型乙酰胆碱受体,现已证明它们是一些化学门控通道,具有能与ACh特异性结合的亚单位。在轴突末梢的轴浆中,除了有许多线粒体外还含有大量直径约50nm的无特殊构造的囊泡(图2-19)。用组织化学的方法可以证明,囊泡内含有ACh;此ACh首先在轴浆中合成,然后贮存在囊泡内。据测定,每个囊泡中贮存的ACh量通常是相当恒定的,且当它们被释放时,也是通过出胞作用,以囊泡为单位“倾囊”释放,被称为量子式释放。在神经末梢处于安静状态时,一般只有少数囊泡随机地进行释放,不能对肌细胞产生显著影响。但当神经末梢处有神经冲动传来时,在动作电位造成的局部膜去极化的影响下,大量囊泡向轴突膜的内侧面靠近,通过囊泡膜与轴突膜的融合,并在融合处出现裂口,使囊泡中的ACh全部进入接头间隙。据推算,一次动作电位的到达,能使大约200~300个囊泡的内容排放,使近107个ACh分子被释放。轴突末梢处的电位变化引起囊泡排放的过程十分复杂,但首先是轴突末梢膜的去极化,引起了该处特有的电压门控式Ca2+通道开放,引起细胞间隙液中的Ca2+进入轴突末梢,触发了囊泡移动以至排放的过程。Ca2+的进入量似乎决定着囊泡释放的数目;细胞外液中低Ca2+或(和)高Mg2+,都可阻碍ACh的释放而影响神经-肌接头的正常功能。已故冯德培院士在30年代对神经-肌接头的化学性质传递进行过重要的研究。
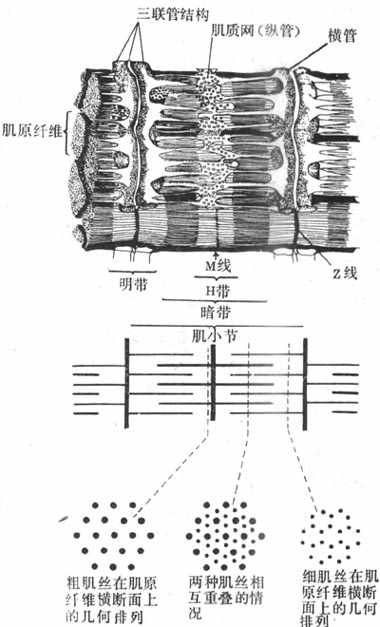
图2-19 神经-肌接头处的超微结构示意图 当ACh分子通过接头间隙到达终板膜表面时,立即同集中存在于该处的特殊通道蛋白质的两个α-亚单位结合,每分子的通道将结合两个分子的ACh,由此引起的蛋白质分子内部构象的变化会导致它的通道结构的开放。这种通道开放时,孔道的横截面比前面提到的Na+通道的面积为大,可允许Na+、K+甚至少量Ca2+同时通过;由于这几种离子正常时在膜内处的分布特点,实际出现的是Na+的内流和K+的外流,其总的结果是使终板膜处原有静息电位减小,向零值靠近,亦即出现膜的去极化;这一电变化,称为终板电位,它的出现约较神经冲动到达接头前膜处晚0.5~1.0ms。有人曾在运动神经无冲动到达末梢时,记录到由于个别囊泡的自发释放在终板膜上引起的微小的电变化,称为微终板电位。终板电位与前述的局部兴奋电反应有类似的性质:不表现“全或无”特性,其大小与接头前膜释放的ACh的量成比例;无不应期,可表现总和现象等,如我们一般记录到的终板电位就是多数微终板电位总和的结果。终板电位产生时,它将以电紧张性扩布的形式影响终板膜周围一般肌细胞膜。一般的肌细胞膜与神经轴突的膜性质类似,其中主要含电压门控式Na+通道和K+通道;因而当同终板膜邻接的肌细胞膜的静息电位由于终板电位的影响而去极化到该处膜的阈电位水平时,就会引发一次向整个肌细胞膜作“全或无”式传导的动作电位,后者再通过所谓“兴奋-收缩耦联”,引起肌细胞出现一次机械收缩。 正常情况下,一次神经冲动所释放的ACh以及它所引起的终板电位的大小,大约超过引起肌细胞膜动作电位所需阈值的3~4倍,因此神经肌接头处的兴奋传递通常是1对1的,亦即运动纤维每有一次神经冲动到达末梢,都能“可靠地”使肌细胞兴奋一次,诱发一次收缩;这一点与将来要讲的神经元之间的兴奋传递有明显不同(见第十章)。接头传递能保持1对1的关系,还要靠每一次神经冲动所释放的ACh能够在它引起一次肌肉兴奋后被迅速清除,否则它将持续作用于终板而使终板膜持续去极化,并影响下次到来的神经冲动的效应。已知,ACh的清除主要靠胆硷酯酶的降解作用来完成,此酶主要分布在接头间隙中和接头后膜上,它们大约可以在2.0ms的时间内将一次神经冲动所释放的ACh清除掉。许多药物可以作用于接头传递过程中的不同阶段,影响正常的接头功能。例如,美洲箭毒和α-银环蛇毒可以同ACh竞争终板膜的ACh受体亚单位,因而可以阻断接头传递而使肌肉失去收缩能力;有类似作用的药物称为肌肉松弛剂;有机磷农药和新斯的明对胆碱酯酶有选择性的抑制作用,可造成ACh在接头和其他部位的大量积聚,引起种种中毒症状。重症肌无力是由于体内骨骼肌终板处的ACh门控通道数量不足或功能障碍所引起。 二、骨骼肌细胞的微细结构 骨骼肌细胞在结构上最突出之点,是含有大量的肌原纤维和丰富的肌管系统,且其排列高度规则有序。肌细胞是体内耗能作功,完成机体多种机械运动的功能单位。 (一)肌原纤维和肌小节 每个肌纤维含有大量直径1~2μm的纤维状结构,称为肌原纤维,它们平行排列,纵贯肌纤维全长,在一个细胞中可达上千条之多(图2-20)。每条肌原纤维的全长都呈现规则的明、暗交替,分别称为明带和暗带;而且在平行的各肌原纤维之间,明带和暗带又都分布在同一水平上;暗带的长度比较固定,不论肌肉处于静止、受到被动牵拉或进行收缩时,它都保持1.5μm的长度;在暗带中央,有一段相对透明的区域,称为H带,它的长度随肌肉所处状态的不同而有变化;在H带中央亦即整个暗带的中央,又有一条横向的暗线,称为M线。明带的长度是可变的,它在肌肉安静时较长,并且在一定范围内可因肌肉所受的被牵引而变长;但明带在肌肉收缩时可变短。明带中央也有一条横向的暗线,称为z线(或z盘)。目前已经肯定,肌原纤维上每一段位于两条z线之间的区域,是肌肉收缩和舒张的最基本单位,它包含一个位于中间部分的暗带和两侧各1/2的明带,合称为肌小节(sarcomere)。由于明带的长度可变,肌小节的长度在不同情况下可变动于1.5~3.5μm之间;通常在体骨骼肌安静时肌小节的长度约为2.0~2.2μm。
图2-20 骨骼肌细胞的肌原纤维和肌管系统 注意肌小节能的组成和不同部位肌小节横断布粗、细肌丝的几何排列 用X-线衍射等更精密的方法进一步发现,肌小节的明带和暗带包含有更细的、平行排列的丝状结构,称为肌丝。暗带中含有的肌丝较粗,直径约10nm,称为粗肌丝,其长度与暗带相同;实际上暗带的形成就是由于粗肌丝的存在,M线则是把成束的粗肌丝固定在一定位置的某种结构。明带中的肌丝较细,直径约5nm,称为细肌丝;它们由z线结构向两侧明带伸出,每侧的长度都是1.0nm,它的游离端在肌小节总长度小于3.5nm的情况下,必然有一段要伸入暗带,和粗肌丝处于交错和重叠的状态;如果由两侧z线伸入暗带的细肌丝未能相遇而隔有一段距离,这就形成了H带。肌肉被拉长时,肌小节长度增大,这时细肌丝由暗带重叠区拉出,使明带长度增大,H带也相应地增大(图2-20,下)。 粗、细肌丝相互重叠时,在空间上也呈现规则排列,这可从肌原纤维的横断面上看出。图2-20下方表示在肌小节的不同位置将肌原纤维横节时,断面上所能看到的两种肌丝的分布情况。在通过明带的横断面上只有细肌丝,它们的所在位置相当于一个正六边形的各顶点;在通过H带的横断面上只有粗肌丝,它们都处于正三边形的各顶点上;而在H带两侧的暗带的横断面上,则可看到粗、细肌丝交错存在的情况,这里,每一条粗肌丝正处在以六条细肌丝为顶点的正六边形的中央,而这就为收缩时粗细肌丝之间的相互作用准备了条件。 (二)肌管系统 肌管系统指包绕在每一条肌原纤维周围的膜性囊管状结构,由来源和功能都不相同的两组独立的管道系统组成。一部分肌管的走行方向和肌原纤维相垂直,称为横管系统或称T管,是由肌细胞的表面膜向内凹入而形成;它们穿行在肌原纤维之间,并在z线水平(有些动物是在暗带和明带衔接处的水平)形成环绕肌原纤维的管道;它们相互交通,管腔通过肌膜凹入处的小孔与细胞外液相通。将标记物加入到细胞的浸浴液中,这些物质可以很快在每一条环绕肌小节的横管系统中出现,但不能进入肌浆和肌浆网中去。肌原纤维周围还有另一组肌管系统,就是肌浆网,它们的走行方向和肌小节平行,称为纵管系统或称为L管;纵管系统或肌浆网主要包绕每个肌小节的中间部分,这是一些相互沟通的管道,但是在接近肌小节两端的横管时管腔出现膨大,称为终末池,它使纵管以较大的面积和横管相靠近。每一横管和来自两侧肌小节的纵管终末池,构成了三联管结构(图2-20,上)。据研究,横管和纵管的膜在三联管结构处并不接触,中间尚隔有约12nm的间隙,说明两组管道的内腔并不直接沟通,但这样的结构显然有利于细胞内外之间某种形式的信息传递。目前普遍承认的看法是,横管系统的作用是将肌细胞兴奋时出现在细胞膜上的电变化沿T管膜传入细胞内部,肌浆网和终末池的作用是通过对钙离子的贮存、释放和再积聚,触发肌小节的收缩和舒张;而三联管结构是把肌细胞膜的电变化和细胞内的收缩过程衔接或耦联起来的关键部位。 三、骨骼肌的收缩机制和兴奋-收缩耦联 Huxley等在50年代初期就提出了用肌小节中粗、细肌丝的相互滑行来说明肌肉收缩的机制。这一被称为滑行理论(sliding theory)的主要内容是:肌肉收缩时虽然在外观上可以看到整个肌肉或肌纤维的缩短,但在肌细胞内并无肌丝或它们所含的分子结构的缩短,而只是在每一个肌小节内发生了细肌丝向粗肌比之间的滑行,亦即由z线发出的细肌丝在某种力量的作用下主动向暗带中央移动,结果各相邻的z线都互相靠近,肌小节长度变短,造成整个肌原纤维、肌细胞乃至整条肌肉长度的缩短。滑行现象最直接的证明是,肌肉收缩时并无暗带长度的变化,而只能看到明带长度的缩短;并且与此同时也看到暗带中央H带相应地变窄。这只能说明,细肌丝在肌肉收缩时也没有缩短,只是它们更向暗带中央移动,和粗肌丝发生了更大程度的重叠。这种变化只能用粗、细肌丝之间出现了相对运动即滑行现象来解释。滑行理论需要进一步说明的问题是:肌肉收缩时究竟是什么力量促使细肌丝向粗肌丝之间滑行,以及怎样把这些过程和肌肉膜的兴奋过程联系起来。近年来,由于肌肉生物化学及其他细胞生物学技术的发展,肌丝滑行的机制已基本上从组成肌丝的蛋白质分子结构的水平得到阐明,对于与滑行的开始和终止有关的各种控制因素,也有了较较深入的了解。 (责任编辑:泉水) |
第四节 肌细胞的收缩功能
时间:2006-06-19 20:38来源:大众医药网 作者:admin 点击:
717次
顶一下
(13)
100%
踩一下
(0)
0%
------分隔线----------------------------
- 发表评论
-
- 最新评论 进入详细评论页>>
